КОРОМЫСЛО ГОЛУБОЕ или СИНЕЕ
Aeshna cyanea Muller, 1764

♂ juv Фото © Виктор Рындевич, Мядельский р-н, Минская обл.
Aeshna cyanea Muller, 1764 (коромысло синее или голубое)
Глаза соприкасаются друг с другом на некотором отрезке. На лбу Т-образный черный рисунок, однако его продольная линия узкая или малозаметная. Тело коричневое, но с крупными зелеными пятнами и полосами. Перед груди бурый с 2 большими удлиненно-овальными желтыми или зелеными пятнами. Доплечевые полосы светло-зеленые, полные и весьма широкие, шире, чем темные полосы между ними. Бока зеленые с черными полосами на швах (средняя прерванная). Сверху голубые пятнышки у основания крыльев. На II сегменте брюшка узкий желтый треугольник (в форме «кубка»); VIII и IX тергиты брюшка сверху с непрерывный голубой или зеленой поперечной полосой. Жилкование крыльев черное. Птеростигма черная, около 3 мм. При посадке принимают вертикальную позу со свисающим вниз брюшком.
♂: Глаза зеленовато-голубые. Сужение брюшка («талия») на III сегменте. Брюшко черное, сверху с зелеными, а по бокам с синими пятнами. 3 последних сегмента брюшка (VIII-X) сверху почти целиком голубые, и пятна по бокам брюшных тергитов также голубые. Вместе с тем, имеется более редкая форма, у которой все сегменты брюшка имеют голубые отметки. Анальный треугольник из трех ячеек. Верхние анальные придатки длинные, с загнутыми книзу концами.
♀: Глаза желтовато-зеленые. Брюшко почти не сужено на III сегменте, коричневато-красное, сверху и по бокам с зелеными пятнами, изредка имеет голубые метки на всех сегментах брюшка. Яйцеклад короткий, задний конец его не заходит даже за конец последнего сегмента брюшка.
У неполовозрелых глаза коричневые, окраска шоколадно-коричневая с бледно-желтыми метками.
Крупная стрекоза, у которой окраска и рисунок пятен на теле сходны с A. juncea, A. mixta и Brachytron pratense. Оба пола A. cyanea можно отличить от трех других видов по наличию двух широких полос на дорсальной поверхности IX и X сегментов брюшка - синей у самцов и зеленой у самок. Доплечевые полосы широкие, в отличие от таковых у A. juncea и A. mixta, у которых они тоньше или вообще отсутствуют. Эти полосы и две широкие полосы по обеим сторонам груди также намного зеленее, чем у A. juncea и A. mixta, у которых они желтые. Более крупные парные пятна на дорсальной поверхности II-VII сегментов брюшка у половозрелых самцов зеленые, а у остальных трех видов - голубые. У A. cyanea грудь не покрыта густыми волосками, как у B. pratense, а птеростигма не такая удлиненная. Вид также намного крупнее.
Длина 67-76 мм, брюшко 51-60 мм, заднее крыло 43-51 мм.
Лет: начало июля - начало октября.
♂: Глаза зеленовато-голубые. Сужение брюшка («талия») на III сегменте. Брюшко черное, сверху с зелеными, а по бокам с синими пятнами. 3 последних сегмента брюшка (VIII-X) сверху почти целиком голубые, и пятна по бокам брюшных тергитов также голубые. Вместе с тем, имеется более редкая форма, у которой все сегменты брюшка имеют голубые отметки. Анальный треугольник из трех ячеек. Верхние анальные придатки длинные, с загнутыми книзу концами.
♀: Глаза желтовато-зеленые. Брюшко почти не сужено на III сегменте, коричневато-красное, сверху и по бокам с зелеными пятнами, изредка имеет голубые метки на всех сегментах брюшка. Яйцеклад короткий, задний конец его не заходит даже за конец последнего сегмента брюшка.
У неполовозрелых глаза коричневые, окраска шоколадно-коричневая с бледно-желтыми метками.
Крупная стрекоза, у которой окраска и рисунок пятен на теле сходны с A. juncea, A. mixta и Brachytron pratense. Оба пола A. cyanea можно отличить от трех других видов по наличию двух широких полос на дорсальной поверхности IX и X сегментов брюшка - синей у самцов и зеленой у самок. Доплечевые полосы широкие, в отличие от таковых у A. juncea и A. mixta, у которых они тоньше или вообще отсутствуют. Эти полосы и две широкие полосы по обеим сторонам груди также намного зеленее, чем у A. juncea и A. mixta, у которых они желтые. Более крупные парные пятна на дорсальной поверхности II-VII сегментов брюшка у половозрелых самцов зеленые, а у остальных трех видов - голубые. У A. cyanea грудь не покрыта густыми волосками, как у B. pratense, а птеростигма не такая удлиненная. Вид также намного крупнее.
Длина 67-76 мм, брюшко 51-60 мм, заднее крыло 43-51 мм.
Лет: начало июля - начало октября.

♀ Фото © Виктор Рындевич, Глубокский р-н, Витебская обл.
Вид обычный, но численность популяции имеет тенденцию к сокращению. Обитает в довольно широком диапазоне типов водоемов, преимущественно на болотных лужах, торфяных выработках, прудах, карьерах, лесных прудах и канавах. Используются также родниковые пруды, медленно текущие ручьи и каналы и особенно садовые и парковые пруды.
По-видимому, нуждается в более низких летних температурах, чем другие виды семейства, поэтому иногда это единственный вид стрекоз, который может оптимально колонизировать небольшие водоемы в лесу, имеющие определенную степень затененности. Тот факт, что небольшие лесные водоемы имеют много валежных структур (упавших в воду веток и стволов) и часто не содержат никакой рыбы, особенно благоприятен для успешного развития личинок.
Имаго взрослеют вдали от воды, встречаются в лесных ландшафтах, часто на лесных полянах, где они охотятся на насекомых размером с бабочку, иногда вместе с другими коромыслами; любят охотиться в полутени, между кустами и под деревьями; теплыми вечерами кормление может продолжаться до заката. Отмечена охота и во время дождя.
Может встречаться совместно с многими другими видами, такими как A. grandis, Anax imperator, Libellula depressa, Sympetrum striolatum. В местах, где водоем занимает совместно с Anax imperator, вид вытесняется на окраины и как таковых собственных территорий не имеет, держась самых заросших участков. Однако пик лёта у Anax imperator кончается несколько раньше и самцы A. cyanea постепенно занимают освободившийся от доминанта водоем.
По-видимому, нуждается в более низких летних температурах, чем другие виды семейства, поэтому иногда это единственный вид стрекоз, который может оптимально колонизировать небольшие водоемы в лесу, имеющие определенную степень затененности. Тот факт, что небольшие лесные водоемы имеют много валежных структур (упавших в воду веток и стволов) и часто не содержат никакой рыбы, особенно благоприятен для успешного развития личинок.
Имаго взрослеют вдали от воды, встречаются в лесных ландшафтах, часто на лесных полянах, где они охотятся на насекомых размером с бабочку, иногда вместе с другими коромыслами; любят охотиться в полутени, между кустами и под деревьями; теплыми вечерами кормление может продолжаться до заката. Отмечена охота и во время дождя.
Может встречаться совместно с многими другими видами, такими как A. grandis, Anax imperator, Libellula depressa, Sympetrum striolatum. В местах, где водоем занимает совместно с Anax imperator, вид вытесняется на окраины и как таковых собственных территорий не имеет, держась самых заросших участков. Однако пик лёта у Anax imperator кончается несколько раньше и самцы A. cyanea постепенно занимают освободившийся от доминанта водоем.

♂ juv Фото © Виктор Рындевич, Шарковщинский р-н, Витебская обл.
Также вид встречается совместно с многими обыкновенными представителями Zygoptera, на которых может иногда охотиться. По крайней мере, случаи охоты отмечены на юго-западе Германии (Баден-Вюртемберг), где лёт вида продолжается дольше, чем в Беларуси, в октябре 2013 г. в послеобеденное время на заболоченном участке наблюдали самца A. cyanea, который неоднократно пытался ловить пары из групп пар Lestes viridis, откладывающих яйца. В течение 30 мин хищник совершил 5 атак, но только дважды сумел схватить самца лютки из тандема. Впрочем, в конце концов во всех случаях пойманные самцы L. viridis сбегали от хищника.
Сначала был схвачен самец лютки, который сразу же отделился от самки. Однако хищник упустил свою добычу, так как жертва вскоре после нападения улетела и не пострадала. Самка L. viridis, разлученная с партнером в результате нападения, вернулась к откладке яиц уже без самца.
При втором нападении самца на тандем L. viridis самец-жертва не отделялся от своей партнерши. Но и этого самца через несколько секунд хищник отпустил и тандем улетел невредимым. В общей сложности за примерно 30 минут наблюдалось еще 3 неудачных нападения на пары L. viridis во время откладки яиц.
На следующий день здесь же наблюдать пары L. viridis, откладывающие яйца, но не группами, как накануне, а отдельными парами. За почти двухчасовой период наблюдения нападения на люток во время яйцекладки не зарегистрировано. Однако в 12:10 одиночный самец L. viridis был пойман самцом A. cyanea, а затем в наземной растительности более крупный хищник съел голову и большую часть груди жертвы.
Является ли L. viridis одной из постоянных жертв A. cyanea или это происходит только в конце сезона, ответ не получен.
Сначала был схвачен самец лютки, который сразу же отделился от самки. Однако хищник упустил свою добычу, так как жертва вскоре после нападения улетела и не пострадала. Самка L. viridis, разлученная с партнером в результате нападения, вернулась к откладке яиц уже без самца.
При втором нападении самца на тандем L. viridis самец-жертва не отделялся от своей партнерши. Но и этого самца через несколько секунд хищник отпустил и тандем улетел невредимым. В общей сложности за примерно 30 минут наблюдалось еще 3 неудачных нападения на пары L. viridis во время откладки яиц.
На следующий день здесь же наблюдать пары L. viridis, откладывающие яйца, но не группами, как накануне, а отдельными парами. За почти двухчасовой период наблюдения нападения на люток во время яйцекладки не зарегистрировано. Однако в 12:10 одиночный самец L. viridis был пойман самцом A. cyanea, а затем в наземной растительности более крупный хищник съел голову и большую часть груди жертвы.
Является ли L. viridis одной из постоянных жертв A. cyanea или это происходит только в конце сезона, ответ не получен.

♀ Фото © Семён Левый, Борисовский р-н, Минская обл.
Самцы территориальны, агрессивно защищают территорию, летая над водой на высоте около 1 м. В Великобритании самая высокая устойчивая плотность составляет около 2 особей на 100 м уреза воды, но в других частях Европы зарегистрированы более высокие плотности. Например, в Швейцарии на пруду 500 м² одновременно наблюдалось до 4-5 самцов. В небольших прудах одновременно может находиться только один самец, хотя из этого водоема из личинок могут отрождаться десятки и даже сотни крылатых особей. Несколько разных самцов занимают один и тот же пруд в течение дня в разное время. Продолжительность каждого визита зависит от количества обитающих в районе: чем их больше, тем более частыми и короткими будут их посещения пруда. Общее время, проведенное у пруда, зависит от температуры воздуха.
Места размножения обычно в нейтральных или щелочных стоячих водах, часто частично затененных деревьями, а иногда и в медленно текущих водах. Встречается в садовых прудах (где на самок, откладывающих яйца, часто охотятся кошки) и может быть единственным видом в тенистых лесных прудах с большим количеством опавших листьев.
Спаривание обычно начинается у воды, хотя может произойти и в местах отдыха на растущих поблизости деревьях. Однако сама копуляция во всех случаях происходит вдали от воды в кронах деревьев и длится до 2 часов.
Яйца самка откладывает обычно выше уровня воды в глинистые берега, гнилые коряги, сырую землю, скопления водных растений, а также в стебли Potamogeton crispus, в моховые подушки Sphagnum sp., в скопления водорослей и т. д. Кладка происходит в августе - сентябре и даже в октябре. Часто она происходит вечером и в плохую погоду. Самка может отложить около 200 яиц в течение часа. Зимуют обычно в фазе яйца.
Места размножения обычно в нейтральных или щелочных стоячих водах, часто частично затененных деревьями, а иногда и в медленно текущих водах. Встречается в садовых прудах (где на самок, откладывающих яйца, часто охотятся кошки) и может быть единственным видом в тенистых лесных прудах с большим количеством опавших листьев.
Спаривание обычно начинается у воды, хотя может произойти и в местах отдыха на растущих поблизости деревьях. Однако сама копуляция во всех случаях происходит вдали от воды в кронах деревьев и длится до 2 часов.
Яйца самка откладывает обычно выше уровня воды в глинистые берега, гнилые коряги, сырую землю, скопления водных растений, а также в стебли Potamogeton crispus, в моховые подушки Sphagnum sp., в скопления водорослей и т. д. Кладка происходит в августе - сентябре и даже в октябре. Часто она происходит вечером и в плохую погоду. Самка может отложить около 200 яиц в течение часа. Зимуют обычно в фазе яйца.

♂ Фото © Виктор Рындевич, Лепельский р-н, Витебская обл.
Нимфа. Личинки от серо-зеленых до серо-желтых, от светло-коричневых до темно-коричневых, почти черных. Окраска личинок, вероятно, находится в зависимости от степени освещенности водоема, прозрачности воды и состава грунта. Особи, находимые в лесных болотах с темным илом, всегда темно-коричневые, почти черные. Из водоемов с чистым дном и открытой хорошо освещаемой солнцем водной поверхностыо личинки обыкновенно светло-коричневые или от серо-зеленых до серо-желтых. Личинки крупные, широкие. Голова уплощенная, суженная сзади, на переднем крае лба светлая линия; темя немного приподнято; глазки хорошо заметны; глаза большие, занимающие полголовы, далеко вдаются назад по направлению к средней линии головы, внутренние задние углы глаз сильно выдающиеся; затылок довольно большой, с сильно волнистым передним краем, вогнутым задним краем, с небольшим приподнятым посредине килем; с каждой стороны от киля по блестящему пятнышку; на верхней стороне задних лопастей несколько блестящих полос, разделенных морщинистыми линиями. Переднегрудь небольшая; внешние края ее острые, светлые, задний край более выпуклый; боковые выросты короткие, тупые, сморщенные, задний шире, тупее и немного длиннее переднего, угол, образуемый ими, немного более чем прямой. Крыловые чехлы доходят до IV сегмента, узкие, светлые. Ноги умеренно тонкие, довольно длинные, с 2 темными четкими кольцами на всех бедрах, с 1-2 такими же кольцами на голенях передней пары ног и с 1 таким же кольцом на остальных голенях. Брюшко массивное, сверху очень выпуклое, гладкое, сильно орнаментированное, расширяющееся постепенно к VI и VII сегментам, затем суживающееся к концу; имеется средняя дорсальная светлая полоса в передней половине каждого сегмента до VIII включительно; по обе стороны от нее проходят две параллельные темные дорсальные полосы, резко обозначенные у начала сегментов; на темных дорсальных полосах вдавления в виде черной поперечной полосы и черной точки позади нее; на каждой стороне большинства сегментов, ближе к внешним краям, по 2 черные точки и по 2 блестящих пятна; IX и Х сегменты орнаментированы менее, чем другие сегменты; темные межсегментальные пространства со светлыми полосами. Дорсальный придаток анальной пирамиды цилиндрический, выемчатый на вершине; церки длинные, острые, треугольные, вершины их часто перекрещиваются; церкоиды с острой тонкой вершиной; основная часть дорсального придатка треугольная с тупой вершиной, равна 1/2 длины церкоидов. Створки яйцеклада очень большие.

♀ Фото © Семён Левый
Личинки - обитатели больших открытых богатых водной растительностью водоемов (озера, пруды, тихие заводи рек и т. д.), а также закрытых лесных болот с илистым дном; встречаются в очень небольших количествах в медленно текущих речках и сточных каналах с богатой водной растительностью, на глубине 0,3-0,5 м. Живут среди водных растений. Нимфы питаются дафниями, личинками комаров и другими водными насекомыми, а на последней стадии своего развития также личинками мелких рыб или амфибий.
Выход личинок из яиц начинается обычно в конце июня или в начале июля следующего года, но иногда личинки вылупляются той же осенью, причем в одном и том же водоеме. Например, в недавно созданном садовом пруду на северо-западе Германии, часть популяции личинок появилась через год после откладки яиц. Другая часть появилась в текущем году и впала в спячку на последних четырех стадиях развитмя. Само развитие личинок обычно длится 2 года, в течение которых происходит до 14 линек. Однако и здесь есть некоторые разночтения. Многочисленные полевые исследования показали, что предличинки вылупляются весной и личинки впадают в зимнюю спячку по крайней мере один раз, прежде чем превратиться в имаго. С другой стороны, имеются данные, указывающие на полный личиночный цикл всего за одно лето. К концу развития они достигают в длину 50 мм. Места окрыления имаго варьируются от высокой маргинальной растительности до деревянных столбов, стволов кустов и даже вертикальных бетонных стен небольших водных резервуаров.
Выход личинок из яиц начинается обычно в конце июня или в начале июля следующего года, но иногда личинки вылупляются той же осенью, причем в одном и том же водоеме. Например, в недавно созданном садовом пруду на северо-западе Германии, часть популяции личинок появилась через год после откладки яиц. Другая часть появилась в текущем году и впала в спячку на последних четырех стадиях развитмя. Само развитие личинок обычно длится 2 года, в течение которых происходит до 14 линек. Однако и здесь есть некоторые разночтения. Многочисленные полевые исследования показали, что предличинки вылупляются весной и личинки впадают в зимнюю спячку по крайней мере один раз, прежде чем превратиться в имаго. С другой стороны, имеются данные, указывающие на полный личиночный цикл всего за одно лето. К концу развития они достигают в длину 50 мм. Места окрыления имаго варьируются от высокой маргинальной растительности до деревянных столбов, стволов кустов и даже вертикальных бетонных стен небольших водных резервуаров.

♀ Фото © Семён Левый
В Великобритании данные за пятилетний период 2007-2011 гг. указывают на бимодальную модель отрождения. Первый, более крупный пик отрождений представлял собой личинок, вступивший в зимовку заблаговременно. Второй, меньший пик состоял из особей, догоняющих в весенний период.
На одном из исследуемых садовых прудов преобладающими растениями, используемыми для отрождения, были сусак и вахта, также использовались болотная незабудка и болотница в каждой локации. Высота отрождения колебалась от 5 до 35 см над уровнем воды, в зависимости от погодных условий, а также вида используемого растения. В теплые дни отрождения, как правило, происходили выше на сусаке. После или во время дождей, как правило, отрождения происходили ниже в менее открытых местах, и предпочтение отдавалось нижней стороне больших широких листьев вахты.
Погодные условия оказывают существенное влияние на метаморфоз личинок. Теплый, солнечный период часто инициирует начало отрождений, особенно если за ним следует влажный и более прохладный период. В оптимальных условиях личинки покидают воду вечером и начинают проходить метаморфоз в 23:00-24:00, имаго готовы к первому полету до рассвета следующего дня. Это дает особям наилучшие шансы завершить отрождение в темное время суток. Когда погода менее благоприятная из-за дождя или холодных ночей, отрождение начинается ранним утром следующего дня.
На одном из исследуемых садовых прудов преобладающими растениями, используемыми для отрождения, были сусак и вахта, также использовались болотная незабудка и болотница в каждой локации. Высота отрождения колебалась от 5 до 35 см над уровнем воды, в зависимости от погодных условий, а также вида используемого растения. В теплые дни отрождения, как правило, происходили выше на сусаке. После или во время дождей, как правило, отрождения происходили ниже в менее открытых местах, и предпочтение отдавалось нижней стороне больших широких листьев вахты.
Погодные условия оказывают существенное влияние на метаморфоз личинок. Теплый, солнечный период часто инициирует начало отрождений, особенно если за ним следует влажный и более прохладный период. В оптимальных условиях личинки покидают воду вечером и начинают проходить метаморфоз в 23:00-24:00, имаго готовы к первому полету до рассвета следующего дня. Это дает особям наилучшие шансы завершить отрождение в темное время суток. Когда погода менее благоприятная из-за дождя или холодных ночей, отрождение начинается ранним утром следующего дня.

♂ Фото © Семён Левый, Кобринский р-н, Брестская обл.
За 5-летний период исследования только дважды было зафиксировано, когда отрождающиеся имаго не смогли освободиться от шкурки личинок. Причины этого не известны. Неспособность расширить и укрепить крылья наблюдалась в трех отдельных случаях и была связана с периодами дождя и сильного ветра. Ветреная и сырая погода повредила двух вылупившихся особей.
Угрозу хищничества для отрождающихся в пруду A. cyanea представляют осы (виды Vespa и Vespula), обыкновенные лягушки, тритоны, воробьи и дрозды. Осы обычно охотятся на только что отродившееся имаго. Лягушки охотятся на молодых имаго, когда те отрождаются на малой высоте. Их привлекает жужжание крыльев, связанное с предполетной обработкой. В то же время, следует отметить, что головастики размножающихся в пруду лягушек в свою очередь являются добычей весенних личинок стрекоз. Тритоны наиболее вероятные хищники на личинок всех возрастов, хотя обычно охоты тритонов на личинок последнего возраста не наблюдалось.
Домовые воробьи, которые размножаются на близлежащих деревьях и в зданиях, были основными видами хищников в пруду, опережая дроздов. Была замечена самка домашнего воробья, поедающая недавно отродившихся A. cyanea. Птица парила, как пустельга, над прудом примерно в 30 см над поверхностью воды. Затем она бросалась, чтобы схватить стрекозу, вылезшую из экзувия, и улететь вместе с ней. Впоследствии это неоднократно наблюдалось в течение года. Эта особь разработала поиск появляющихся стрекоз и использовала стратегию добычи пищи. Такого поведения не наблюдалось ни в один другой год.
Угрозу хищничества для отрождающихся в пруду A. cyanea представляют осы (виды Vespa и Vespula), обыкновенные лягушки, тритоны, воробьи и дрозды. Осы обычно охотятся на только что отродившееся имаго. Лягушки охотятся на молодых имаго, когда те отрождаются на малой высоте. Их привлекает жужжание крыльев, связанное с предполетной обработкой. В то же время, следует отметить, что головастики размножающихся в пруду лягушек в свою очередь являются добычей весенних личинок стрекоз. Тритоны наиболее вероятные хищники на личинок всех возрастов, хотя обычно охоты тритонов на личинок последнего возраста не наблюдалось.
Домовые воробьи, которые размножаются на близлежащих деревьях и в зданиях, были основными видами хищников в пруду, опережая дроздов. Была замечена самка домашнего воробья, поедающая недавно отродившихся A. cyanea. Птица парила, как пустельга, над прудом примерно в 30 см над поверхностью воды. Затем она бросалась, чтобы схватить стрекозу, вылезшую из экзувия, и улететь вместе с ней. Впоследствии это неоднократно наблюдалось в течение года. Эта особь разработала поиск появляющихся стрекоз и использовала стратегию добычи пищи. Такого поведения не наблюдалось ни в один другой год.

♂ Фото © Семён Левый, Житковичский р-н, Гомельская обл.

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♂ Image © Lisa Sundström / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
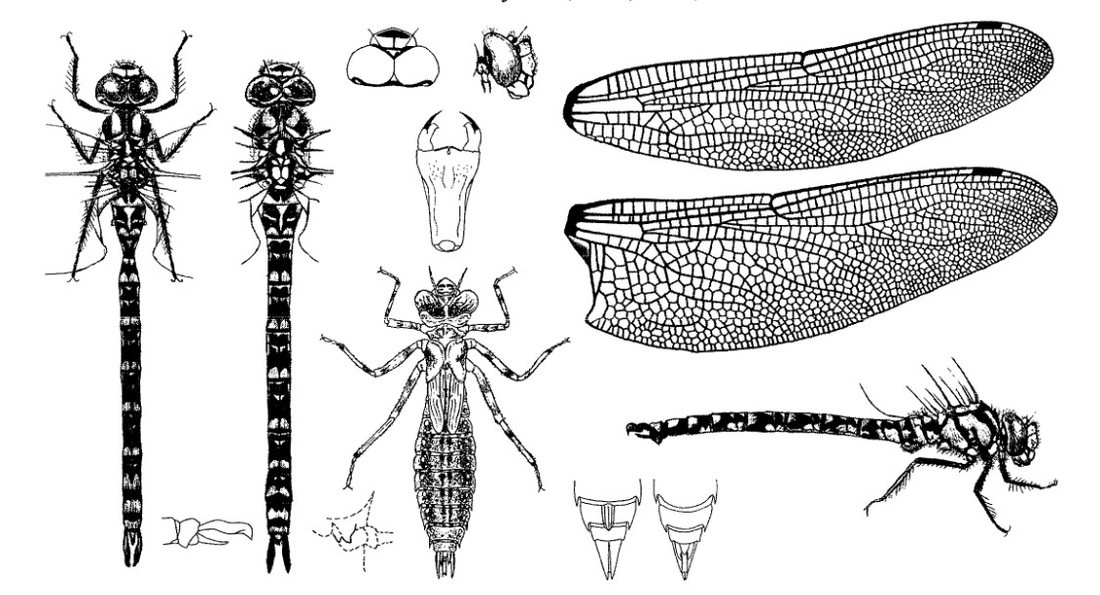
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
7. Moreno-Benítez J. M., Ripoll J. "Aeshna cyanea" / in: Dragonflies along the Great Málaga Path and in the Province. Identification Guide, 2018. pp. 96-97.
8. Jodicke R. "Nachweis einjähriger Entwicklung bei Aeshna cyanea (Müller) (Anisoptera: Aeshnidae)" / Libellula 18 (3/4): 169-174. 1999.
9. Schneider B., Wildermith H. "Libellen als Individuen – zum Beispiel Aeshna cyanea (Odonata: Aeshnidae)" / Entomo Helvetica, 2: 185-199, 2009.
10. Smallshire D., Swash A. "Southern Hawker Aeshna cyanea" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 110-111.
11. Smallshire D., Swash A. "Blue Hawker. Aeshna cyanea" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 142-143.
12. Nowak M. "Blaugrüne Mosaikjungfer Aeshna cyanea auf systematischer Jagd nach Tandems der Weidenjungfer Lestes viridis" / Mercuriale - Libellllen in Baden-Württemberg. Band 13. 2013. 41-42.
13. Cham S. "A study of Southern Hawker Aeshna cyanea emergence from a garden pond" / Journal of the British Dragonfly Society, Volume 28 No. 1, 2012. pp. 1-20.
14. Mauersberger R., Brauner O. "Blaugrüne Mosaikjungfer – Aeshna cyanea (Müller 1764)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 66-67.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
7. Moreno-Benítez J. M., Ripoll J. "Aeshna cyanea" / in: Dragonflies along the Great Málaga Path and in the Province. Identification Guide, 2018. pp. 96-97.
8. Jodicke R. "Nachweis einjähriger Entwicklung bei Aeshna cyanea (Müller) (Anisoptera: Aeshnidae)" / Libellula 18 (3/4): 169-174. 1999.
9. Schneider B., Wildermith H. "Libellen als Individuen – zum Beispiel Aeshna cyanea (Odonata: Aeshnidae)" / Entomo Helvetica, 2: 185-199, 2009.
10. Smallshire D., Swash A. "Southern Hawker Aeshna cyanea" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 110-111.
11. Smallshire D., Swash A. "Blue Hawker. Aeshna cyanea" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 142-143.
12. Nowak M. "Blaugrüne Mosaikjungfer Aeshna cyanea auf systematischer Jagd nach Tandems der Weidenjungfer Lestes viridis" / Mercuriale - Libellllen in Baden-Württemberg. Band 13. 2013. 41-42.
13. Cham S. "A study of Southern Hawker Aeshna cyanea emergence from a garden pond" / Journal of the British Dragonfly Society, Volume 28 No. 1, 2012. pp. 1-20.
14. Mauersberger R., Brauner O. "Blaugrüne Mosaikjungfer – Aeshna cyanea (Müller 1764)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 66-67.
