КОРОМЫСЛО БОЛЬШОЕ
Aeshna grandis Linnaeus, 1758

♂ Фото © Виктор Рындевич, Минский р-н
Aeshna grandis Linnaeus, 1758 (коромысло большое)
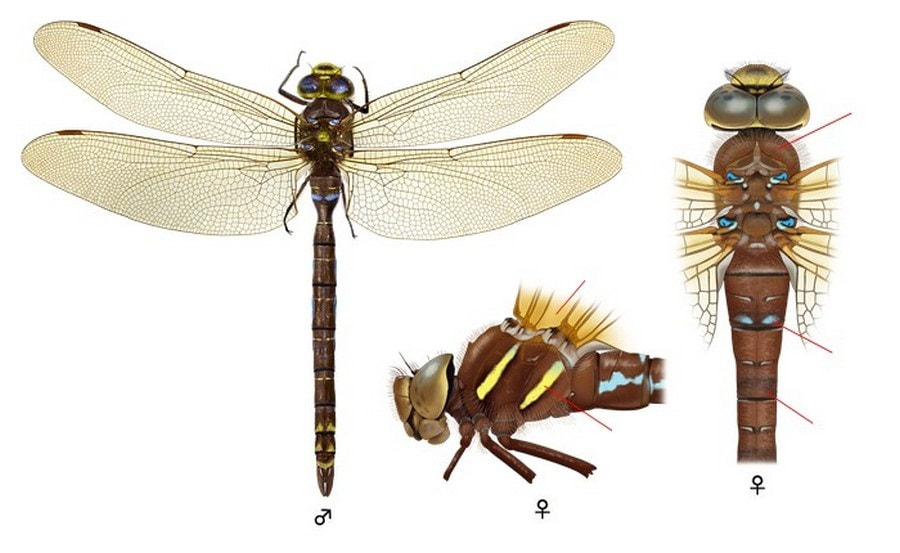
Легко отличается от других видов рода красновато-коричневой окраской тела, дымчатыми буроватыми крыльями, бурыми жилками и полным отсутствием доплечевых полос. Глаза соприкасаются друг с другом на длинном отрезке. Лоб с бледно-бурым Т-образным пятном, без продольной черты или она очень тонкая и неясно выраженная. Шов между лбом и наличником не зачернен. Тело сверху однотонно-коричневое, но с 4 хорошо заметными голубыми пятнышками у основания крыльев. Бока груди коричневые с 2 четко видными желтоватыми или желто-зелеными полосами. Доплечевые полосы отсутствуют. Жилкование крыльев рыже-коричневое. Крылья равномерно окрашены в янтарный цвет. Птеростигма белая. При посадке принимают вертикальную позу со свисающим вниз брюшком. Ноги сплошь рыжие или красно-бурые.
♂: Глаза коричневые, но сверху голубые. Основная окраска равномерно-коричневая. По бокам груди две неширокие, но четко выделяемые желтые полосы. Брюшко коричневое или темно-бурое по бокам. На дорсальной поверхности II сегмента брюшка пара темно-голубых или синих пятен, на III сегменте сбоку большая отметка такого же цвета, по бокам IV-VIII сегментов более мелкие синие метки. На дорсальной поверхности II-VIII сегментов брюшка крошечные парные желтые отметины.
♀: Глаза желтовато-коричневые. Брюшко коричневое или темно-бурое с очень маленькими желтыми пятнышками. 3-е брюшное кольцо не сужено. Окраска как у самца, но голубые пятна на II сегменте отсутствуют (иногда редуцированы), а отметины по бокам брюшка желтые. Однако у некоторых самок имеются голубые метки по бокам брюшка. Яйцеклад короткий, задний конец его не заходит даже за конец последнего сегмента брюшка.
Неполовозрелые окрашены как самки.
Лет: конец июня - начало октября.
Длина 70-77 мм, брюшко 49-60 мм, заднее крыло 45-49 мм.
♂: Глаза коричневые, но сверху голубые. Основная окраска равномерно-коричневая. По бокам груди две неширокие, но четко выделяемые желтые полосы. Брюшко коричневое или темно-бурое по бокам. На дорсальной поверхности II сегмента брюшка пара темно-голубых или синих пятен, на III сегменте сбоку большая отметка такого же цвета, по бокам IV-VIII сегментов более мелкие синие метки. На дорсальной поверхности II-VIII сегментов брюшка крошечные парные желтые отметины.
♀: Глаза желтовато-коричневые. Брюшко коричневое или темно-бурое с очень маленькими желтыми пятнышками. 3-е брюшное кольцо не сужено. Окраска как у самца, но голубые пятна на II сегменте отсутствуют (иногда редуцированы), а отметины по бокам брюшка желтые. Однако у некоторых самок имеются голубые метки по бокам брюшка. Яйцеклад короткий, задний конец его не заходит даже за конец последнего сегмента брюшка.
Неполовозрелые окрашены как самки.
Лет: конец июня - начало октября.
Длина 70-77 мм, брюшко 49-60 мм, заднее крыло 45-49 мм.

♀ Фото © Виктор Рындевич, Смолевичский р-н, Минская обл.
Вид обычный. Численность характеризуется либо заметным сокращением, либо довольно быстрым увеличением особей популяции.
Типичный вид лесных и озерных территорий. Встречается вблизи или около рек и различных водоемов: озер, прудов, болот, стариц рек. Предпочтение отдается водоемам со стоячей водой или очень медленным водотоком. На крупных озерах заселяет богатые растительностью заливы или участки с обширными зонами заиления. Реже селится в прудах, каналах, канавах и в болотах. В то же время, для Великобритании указывается как один из видов, наиболее часто встречающихся крупных стрекоз, часто посещающая садовые пруды. По-видимому, вид не привязан к определенным структурам растительности, но явно предпочитает водоемы в лесу или рядом с ним. Кроме того, предпочтение отдается зрелым водам с богато структурированной тростниковой зоной. Также регулярно встречается в кислых водах, таких как болотные пруды и болотные озера. На проточных водоемах, откуда поступает около 10% свидетельств, заселяет слаботочные, часто заболоченные участки, но в большинстве локаций можно найти лишь несколько экзувий на водоем. Поэтому можно предположить, что целый комплекс более или менее соседних водоемов используется для воспроизводства одной популяцией в смысле понятия метапопуляции.
Типичный вид лесных и озерных территорий. Встречается вблизи или около рек и различных водоемов: озер, прудов, болот, стариц рек. Предпочтение отдается водоемам со стоячей водой или очень медленным водотоком. На крупных озерах заселяет богатые растительностью заливы или участки с обширными зонами заиления. Реже селится в прудах, каналах, канавах и в болотах. В то же время, для Великобритании указывается как один из видов, наиболее часто встречающихся крупных стрекоз, часто посещающая садовые пруды. По-видимому, вид не привязан к определенным структурам растительности, но явно предпочитает водоемы в лесу или рядом с ним. Кроме того, предпочтение отдается зрелым водам с богато структурированной тростниковой зоной. Также регулярно встречается в кислых водах, таких как болотные пруды и болотные озера. На проточных водоемах, откуда поступает около 10% свидетельств, заселяет слаботочные, часто заболоченные участки, но в большинстве локаций можно найти лишь несколько экзувий на водоем. Поэтому можно предположить, что целый комплекс более или менее соседних водоемов используется для воспроизводства одной популяцией в смысле понятия метапопуляции.

♂ Фото © Инна Макаревич, Новогрудский р-н, Гродненская обл.
Патрулируют водоемы и прилегающую надводную растительность, но часто находятся вдали от воды, иногда большими группами. В безветренную погоду часто летают на уровне кроны деревьев и активны до позднего вечера. Имаго часто можно наблюдать охотящимся на лесных опушках и полянах, защищенных от ветра лугах. Регулярно охотится в ночное время и остается активным даже при низких температурах (до 12°C), высокой влажности (до 87%) и слабом освещении (до 5 лк). Многие авторы указывают на его вечернюю кормежку, а некоторые считают единственной стрекозой в Центральной Европе, которая добывает пищу даже после полуночи.
Полет характерный, включает в себя длительное планирование, чередующееся с быстрыми, но довольно мелкими взмахами крыльев. Исключительно искусны в полете и способны к внезапным скачкам скорости в очень ограниченном пространстве или изменению направления, летая вертикально, боком или назад, или зависая на месте. Добыча, в том числе комары, мухи и другие крылатые насекомые, ловится в полете благодаря широкому спектру зрения, который стрекоза получает, поворачивая голову на угол до 180° и используя огромные полусферические глаза. Нередко ненадолго садятся чтобы съесть добычу, но в целом виду свойственны продолжительные полеты на большой высоте без посадок.
Отмечаются разлеты на дальние расстояния от водоемов. Из-за такого миграционного поведения вид может появляться в больших количествах в районах, где обычно встречается редко или вообще отсутствует.
Полет характерный, включает в себя длительное планирование, чередующееся с быстрыми, но довольно мелкими взмахами крыльев. Исключительно искусны в полете и способны к внезапным скачкам скорости в очень ограниченном пространстве или изменению направления, летая вертикально, боком или назад, или зависая на месте. Добыча, в том числе комары, мухи и другие крылатые насекомые, ловится в полете благодаря широкому спектру зрения, который стрекоза получает, поворачивая голову на угол до 180° и используя огромные полусферические глаза. Нередко ненадолго садятся чтобы съесть добычу, но в целом виду свойственны продолжительные полеты на большой высоте без посадок.
Отмечаются разлеты на дальние расстояния от водоемов. Из-за такого миграционного поведения вид может появляться в больших количествах в районах, где обычно встречается редко или вообще отсутствует.

♀ Фото © Николай Гурков, г. Витебск
Местами, используемыми для размножения и развития личинок, являются преимущественно стоячие и слабопроточные воды, нейтральные или кислые, на открытых или лесных участках: пруды старые карьеры, торфяники, озера, тихие участки рек и каналов, заводи. Вид нуждается в воде от мезотрофной до эвтрофной, и ему хорошо подходят пруды с полным или частичным пересыханием летом, в дополнение к другим средам обитания, упомянутым выше. В любом случае, болота и лесные пруды кажутся наиболее благоприятными для вида. Также упоминается, что эти места должны по крайней мере включать участки лесистых берегов. Встречается совместно со многими другими видами стрекоз, которые размножаются в прудах и водоемах с хорошей растительностью.
Вид считается не особо территориальным, но в некоторых местах, например, в Британии плотность взрослых самцов редко превышает 2 особи на 100 м уреза воды. Необычна также свойственная этому виду защита территорий вдали от воды. Вместе с тем, в отличие от других коромысел, самцы во время размножения конфликтуют нечасто.
Вид считается не особо территориальным, но в некоторых местах, например, в Британии плотность взрослых самцов редко превышает 2 особи на 100 м уреза воды. Необычна также свойственная этому виду защита территорий вдали от воды. Вместе с тем, в отличие от других коромысел, самцы во время размножения конфликтуют нечасто.

♂ Фото © Татьяна Алешка, Лепельский р-н, Витебская обл.
Несмотря на широкую распространенность вида долгое время было удивительным почти полное отсутствие данных, касающихся брачного поведения - где, когда и как самцы этого вида встречаются со своими самками. Поэтому небезынтересны исследования в обширных заболоченных пастбищах с сетью дренажных канав на северо-западе Германии в Нижней Саксонии. Обнаружено, что спаривание в основном происходит на рассвете. С первыми утренними лучами самцы начинают поиски самок, которые укрылись глубоко в наземной растительности, состоящей из тростника, камыша и травы. Задокументировано утреннее спаривание в характерном положении скрученного кольца. Эти кольца уникальны тем, что образуются не в полете, а сидя на растительности. Перекрученное положение, когда самец висит вниз головой, а его придатки наклонно скользят по голове самки, является результатом образования брачных колес с сидящей самкой. Ближе к утру наблюдали кормовой полет в подходящих местах и отдых в низкой растительности сырого луга. Во время фазы отдыха некоторые самцы осматривали растительность в так называемом «утреннем поисковом полете». В это время, а также при аггрегации кормящихся особей обнаружены раскрученные, вертикально висящие пары, которые были интерпретированы как кольца, образованные в полете, что указывало на альтернативную тактику спаривания.

♂ Фото © Инна Макаревич, Новогрудский р-н, Гродненская обл.
Самцы регулярно совершали утренний поисковый полет до и во время восхода солнца. Ранние утренние пары спаривались в позе скрученного колеса и никогда не пытались сбежать, когда к ним приближался наблюдатель. Местами встреч самцов и самок были густые заросли травы и камыша на нескошенных влажных лугах, тростниковые полосы вдоль канав и заросли Phragmites australis и Glyceria maxima на краях влажных лугов. Численность ищущих самцов различалась в зависимости от дня, места наблюдений и погодных условий на рассвете.
В период между 33 и 14 минутами до восхода солнца замечены первые особи, летающие вокруг растительности. Несколько особей охотились на комаров, но большинство совершало поисковые полеты. Они пролетали низко над густой растительностью, но время от времени глубоко проникали в нее. Даже в тростниковых насаждениях высотой 2 м они осматривали субстрат близко к земле. В росистое утро такие "заныривания" делали их очень мокрыми, поэтому время от времени самцам приходилось вылезать из растительности, чтобы стряхнуть с себя воду. При этом они совершали быстрые вращения вниз, включая три-четыре сальто, чтобы избавиться от капель росы. Отмечались только ищущие самцы, которые встречаясь друг с другом не вели себя агрессивно. Неоднократно наблюдалось, как ищущие самцы прекращали летать, усаживались в растительности и вяло отдыхали. Поисковые действия закончились между 4 и 35 минутами после восхода солнца. Только при сильной облачности и ветре поиски продолжались до 47 мин после восхода солнца. Период поискового полета на рассвете длился 28-63 мин (в среднем 45±11).
В период между 33 и 14 минутами до восхода солнца замечены первые особи, летающие вокруг растительности. Несколько особей охотились на комаров, но большинство совершало поисковые полеты. Они пролетали низко над густой растительностью, но время от времени глубоко проникали в нее. Даже в тростниковых насаждениях высотой 2 м они осматривали субстрат близко к земле. В росистое утро такие "заныривания" делали их очень мокрыми, поэтому время от времени самцам приходилось вылезать из растительности, чтобы стряхнуть с себя воду. При этом они совершали быстрые вращения вниз, включая три-четыре сальто, чтобы избавиться от капель росы. Отмечались только ищущие самцы, которые встречаясь друг с другом не вели себя агрессивно. Неоднократно наблюдалось, как ищущие самцы прекращали летать, усаживались в растительности и вяло отдыхали. Поисковые действия закончились между 4 и 35 минутами после восхода солнца. Только при сильной облачности и ветре поиски продолжались до 47 мин после восхода солнца. Период поискового полета на рассвете длился 28-63 мин (в среднем 45±11).

♂ Фото © Семён Левый, Смолевичский р-н, Минская обл.
В утренние часы отмечено несколько самок, отдыхающих в густой растительности, над которой совершался поисковый полет. Они сидели у земли, на высоте 10-50 см, в более или менее вертикальном висячем положении. Их положение сохранялось до 03:25 после восхода солнца. После дефекации и чистки самки без предварительного жужжания взлетели и покинули место.
Также обнаружено одно искривленное сопрягающееся кольцо в густой растительности. Самец находился в типичном висячем положении, голова была обращена вниз, а кончик брюшка соскользнул за левый глаз самки. Пара оказалась в неловкой ситуации, так как самка цеплялась за тростниковый лист всего тремя коготками двух лапок, а самец не имел контакта ни с какой опорой, кроме своей самки. Эта пара в таком положении была обнаружена через 39 мин после восхода солнца и через 35 мин после регистрации последнего ищущего самца. На этом этапе пара была неактивна, но еще через 25 мин были отмечены ритмичные копулятивные движения. Через 90 мин, когда самка цеплялась лишь одной лапкой за лист тростника, исследователи помогли паре и переместили ее так, чтобы самец смог обхватить стебель тростника. Ровно через три часа после восхода солнца пара прекратила совокупление. Сначала было замечено два резких взмаха крыльев самца вверх, особенно передних. Затем пара разъединила генитальную связку, образовав посткопуляционный тандем на несколько секунд, прежде чем самец ослабил хватку. В этом коротком положении самка могла хорошо зацепиться за стебель. Через несколько секунд она ненадолго взлетела и снова укрылась в растительности. Через минуту самец тоже взлетел и тут же начал новый поисковый полет.
Также обнаружено одно искривленное сопрягающееся кольцо в густой растительности. Самец находился в типичном висячем положении, голова была обращена вниз, а кончик брюшка соскользнул за левый глаз самки. Пара оказалась в неловкой ситуации, так как самка цеплялась за тростниковый лист всего тремя коготками двух лапок, а самец не имел контакта ни с какой опорой, кроме своей самки. Эта пара в таком положении была обнаружена через 39 мин после восхода солнца и через 35 мин после регистрации последнего ищущего самца. На этом этапе пара была неактивна, но еще через 25 мин были отмечены ритмичные копулятивные движения. Через 90 мин, когда самка цеплялась лишь одной лапкой за лист тростника, исследователи помогли паре и переместили ее так, чтобы самец смог обхватить стебель тростника. Ровно через три часа после восхода солнца пара прекратила совокупление. Сначала было замечено два резких взмаха крыльев самца вверх, особенно передних. Затем пара разъединила генитальную связку, образовав посткопуляционный тандем на несколько секунд, прежде чем самец ослабил хватку. В этом коротком положении самка могла хорошо зацепиться за стебель. Через несколько секунд она ненадолго взлетела и снова укрылась в растительности. Через минуту самец тоже взлетел и тут же начал новый поисковый полет.

♀ Фото © Николай Гурков, г. Витебск
Утром, после окончания поисковых полетов, вдоль канав отмечалось самцов меньше, чем самок. Самцы отдыхали глубоко в растительности, близко к земле. Как только эти участки осветило солнце, самцы ориентировались спиной к нему, прогревались и снова начали летать. Их видели с 08:43 и позже (3 ч после восхода солнца и 5 ч до полудня) патрулирующими вдоль канав, иногда кормящихся и осматривающих прибрежную растительность. Самки оставались в позе покоя до тех пор, пока полностью не прогрелись солнцем, а затем покидали участок.
Ситуация утром на двух других локациях была более сложной. На заключительном этапе поисковых полетов все самцы покинули место встреч и улетели над деревьями в сторону берега озера и прилегающих мест. В первые два часа после поисковых полетов в этих локациях практически не было замечено ни одной особи, за исключением нескольких одиночных самцов и самок, которые все еще отдыхали глубоко в растительности. На третий час все больше и больше особей, как самцов, так и самок, оседали на лугу, и, соответственно, было видно, как они летели над деревьями от озера, как правило, в охотничьем полете. Отдыхающие особи были очень пугливы, но, когда их тревожили, их бегство было кратким (около 10-20 м), прежде чем они снова приземлялись в траве или в камыше.
Утром самцы летали низко над лугом и осматривали места с густой растительностью. Этот полет напоминал утренний поисковый полет, но был более прямым, быстрым и с менее тщательным осмотром зарослей.
Ситуация утром на двух других локациях была более сложной. На заключительном этапе поисковых полетов все самцы покинули место встреч и улетели над деревьями в сторону берега озера и прилегающих мест. В первые два часа после поисковых полетов в этих локациях практически не было замечено ни одной особи, за исключением нескольких одиночных самцов и самок, которые все еще отдыхали глубоко в растительности. На третий час все больше и больше особей, как самцов, так и самок, оседали на лугу, и, соответственно, было видно, как они летели над деревьями от озера, как правило, в охотничьем полете. Отдыхающие особи были очень пугливы, но, когда их тревожили, их бегство было кратким (около 10-20 м), прежде чем они снова приземлялись в траве или в камыше.
Утром самцы летали низко над лугом и осматривали места с густой растительностью. Этот полет напоминал утренний поисковый полет, но был более прямым, быстрым и с менее тщательным осмотром зарослей.

♀ Фото © Виталий Сирота, Глубокский р-н, Витебская обл.
В еще одной локации отмечено около 50 кормящихся особей в течение 90 мин в период до и около полудня. Этот участок был защищен от порывистого ветра зарослями ольхи и ивы. Aeshna grandis сопровождали A. cyanea и A. viridis; все охотились на комаров и более крупных неопознанных насекомых, которых поедали, сидя на внешних ветвях периферийных кустов. Высота полета кормящихся особей составляла 2-4 м, иногда до 6 м, что соответствовало высоте деревьев по участкам тростника. Остальные особи, как самцы, так и самки, отдыхали на верхних побегах тростника. Между кормящимися особями не отмечено агрессии, но самки регулярно демонстрировали отказ, изгибая вниз брюшко при приближении самца.

♂ Фото © Владимир Бондарь, Могилевский р-н
В целом, все европейские представители Aeshnidae обычно начинают копуляцию в воздухе, поскольку им нужен полет для сформирования брачного кольца сразу после создания прекопулятивного тандема. Затем совокупление продолжается на растительности в висячем положении. Ищущие самцы A. grandis и A. viridis приближаются к сидящим самкам только рано утром и образуют так называемое «скрученное кольцо», в результате чего самка сохраняет свое сидячее положение. Все скрученные кольца можно узнать по кончику брюшка самца, скользящему по одному из глаз самки в косом положении. Само приближения самца к сидящей восприимчивой самке не наблюдалось, но можно сделать вывод из скрученного положения, что самец прижимает свои анальные придатки к голове самки, а затем опускает все тело, чтобы достичь генитального сцепления.
В конечном положении самец висит под своей партнершей вниз головой, иногда цепляясь за тростник, а иногда нет. Последующее совокупление может длиться часами.
В конечном положении самец висит под своей партнершей вниз головой, иногда цепляясь за тростник, а иногда нет. Последующее совокупление может длиться часами.

♂ Фото © Владимир Бондарь, Могилевский р-н
Активность поздним утром не следует строгому графику, а зависит от погодных условий и физического состояния особей. Большинство самцов покидают место встречи с самками после утреннего поискового полета и летят к подходящим для кормления местам, как и самки, как только места их ночлега полностью прогреваются солнцем. В солнечные дни самцы начинают возвращаться в канавы примерно через 3,5 ч после восхода солнца, где совершают патрульные полеты. Самки следуют туда примерно через 2 часа для откладки яиц.
Типичным элементом поведения в середине утра, по-видимому, является период отдыха после кормового полета. Ежедневно наблюдалось, как вид охотится в болотистых лесах, а затем оседает в низкой растительности. Пары, замеченные в середине утра на местах отдыха и кормежки, отличались от раннеутренних скрученных колец тем, что летели легко и не находились в скрученном положении. Эти дневные кольца образовались в полете (воздушная инициация), как описывалось ранее для вида. Оказалось, что восприимчивые самки используют альтернативную тактику для встречи с самцами, что можно интерпретировать как второй шанс, если они не были обнаружены и оплодотворены на рассвете. Однако основной и наиболее важной тактикой спаривания является утренняя.
Типичным элементом поведения в середине утра, по-видимому, является период отдыха после кормового полета. Ежедневно наблюдалось, как вид охотится в болотистых лесах, а затем оседает в низкой растительности. Пары, замеченные в середине утра на местах отдыха и кормежки, отличались от раннеутренних скрученных колец тем, что летели легко и не находились в скрученном положении. Эти дневные кольца образовались в полете (воздушная инициация), как описывалось ранее для вида. Оказалось, что восприимчивые самки используют альтернативную тактику для встречи с самцами, что можно интерпретировать как второй шанс, если они не были обнаружены и оплодотворены на рассвете. Однако основной и наиболее важной тактикой спаривания является утренняя.

♀ Фото © Виталий Сирота, Глубокский р-н, Витебская обл.
Как уже указывалось, спаривание довольно продолжительное, самки впоследствии откладывают яйца в одиночку. Яйца откладываются либо в живой, либо в мертвый растительный материал, вблизи или ниже поверхности воды. Для кладки используются отмершие ткани растений, погруженные в воду, или скопления отмерших частей растений у берегов водоемов, а также нижняя сторона листьев кубышек и кувшинок. Особенно охотно самки кладут яйца в погруженную в воду древесину. Самка при кладке опускает в воду брюшко только до крыльев, не далее. Яйца откладываются иногда в сырую землю и для кладки предпочтительно выбираются тенистые места среди разросшихся по берегам кустарников, стеблей осоки, под корягами и т. д. Иногда несколько самок группами обосновываются на небольшом плавающем бревне и откладывают яйца близко друг к другу, но в других случаях они проявляют агрессивность по отношению друг к другу. Яйца зимуют в диапаузе. Развитие яиц продолжается около 9 месяцев в связи с уходом на зимовку.

♀ Фото © Виктор Рындевич, Смолевичский р-н, Минская обл.
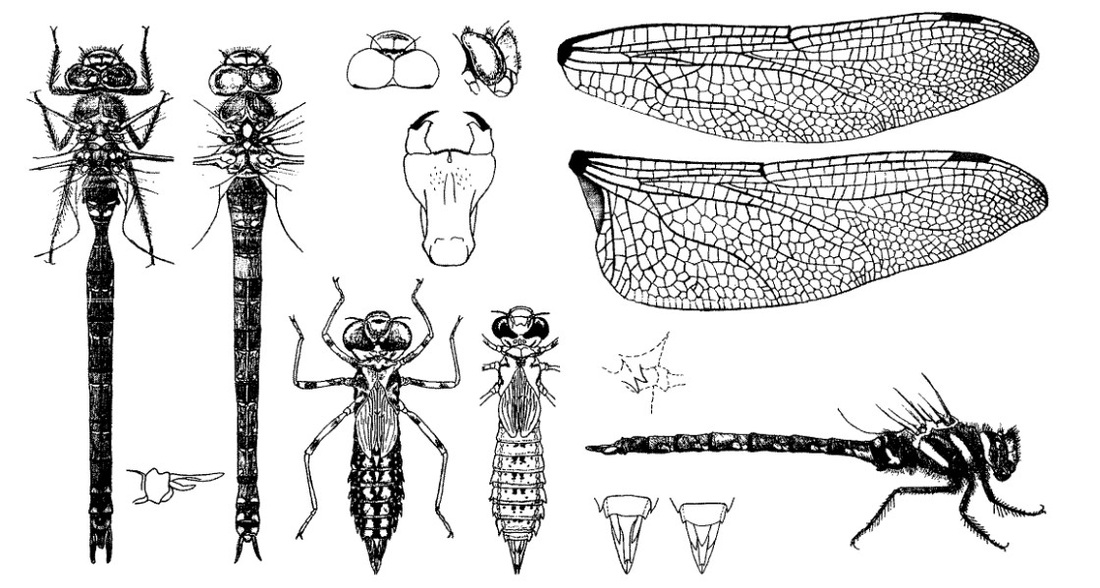
Нимфа. Две формы, отличающиеся по окраске тела, светлая и темная; последняя преобладает. Основная окраска светлых форм однотонная, соломенно-желтая, со светло-коричневыми слабыми отметинами на дорсальной стороне брюшка; темных - коричневая, варьирующая в оттенках у разных особей от светло-коричневой до темно-коричневой, почти черной; обе формы, особенно темная, пестро разрисованные; тело продолговатое, суженное позади груди. Голова плоская, довольно широкая; глаза большие, грушевидные, с заостренным продолжением назад по направлению к срединной линии головы; затылок относительно широкий, с извилистым передним краем, с большой неглубокой выемкой на заднем крае, с 2 светлыми большими продольными полосами на округлых задних лопастях. Переднегрудь с коротким приподнятым передним краем и длинным задним, с сильно скошенными боковыми сторонами, с острыми светлыми углами; боковые выросты переднегруди острые, с вершинами, направленными в разные стороны, задний вырост длиннее и шире переднего. Крыловые чехлы широкие. Ноги довольно длинные, толстые, красноватые или коричневые, у светлых форм почти без рисунка; у темных на бедрах всех ног по 3 светлые поперечные полосы. Брюшко постепенно расширяющееся к VII сегменту, затем суживающееся к концу, пестро разрисованное, у различных особей по окраске варьирующее. В то время как рисунки головы и груди весьма постоянны, рисунки брюшка сильно варьируют. Пятнистая форма: срединные вдавления в виде поперечных коротких черт и точек, а также большое округлое срединное пятно у заднего края сегмента темные; светлая срединная полоса напоминает собою трехзубый листок; обе светлые широкие продольные полосы имеются только на передних сегментах, а начиная с VI сегмента они редуцируются в пятна неопределенной формы. Полосатая форма: посредине через все брюшко проходит коричневая или черновато-коричневая широкая (около 2 мм) продольная полоса (срединная полоса), на VIII-Х сегментах или только на VIII и IX сплошная, а на IV-VII сегментах посредине со светлой продолговатой полоской, занимающей около 1/2 длины сегмента; срединные вдавления и основное пятно лежат на этой полосе; по обе стороны от срединной полосы тянутся 2 светлые широкие продольные полосы с незначительным боковым расширением (срединно-боковые полосы); на каждом сегменте, за исключением Х сегмента, близ внешних краев находятся светлые вдавления или в виде одной полоски, или в виде двух кружков, напоминающих цифру 8, занимающие верхние 2/3 сегментов. Промежуточная форма: средняя темная продольная полоса не сплошная, а в виде пятен, отсутствующих на IX и Х сегментах. У промежуточных форм светлые полосы широкие.
Анальная пирамида длинная; остроконечные длинные церки длиннее дорсального придатка; церкоиды острые, цилиндрические; основная часть дорсального придатка треугольная, тупозаостренная. Створки яйцеклада самки не достигают заднего края IX стернита, длина их 2,5 мм.
Анальная пирамида длинная; остроконечные длинные церки длиннее дорсального придатка; церкоиды острые, цилиндрические; основная часть дорсального придатка треугольная, тупозаостренная. Створки яйцеклада самки не достигают заднего края IX стернита, длина их 2,5 мм.

♀ Фото © Николай Гурков, г. Витебск
Личинки достигают в длину 50 мм, развиваются 2-3 года (по другим данным, 14-26 месяцев, но есть и указание французских авторов, что в зависимости от региона развитие может достигать 5 лет). Предпочитают стоячие или слабопроточные водоемы. Переносят умеренные уровни загрязнения. Населяют самые разнообразные водоемы: ямы, болота, пруды, озера, речки и реки со слабым течением; живут обыкновенно среди полусгнивших веток и стеблей растений, коряг, погруженных в воду бревен, щепок, палок, дров и т. п., на глубине 0,1-1,0 м.
Окрыление происходит на растениях, растущих около берега на высоте от 10 см до 1 м, однако окончательное формирование имаго наблюдается на кустарниках и деревьях. Вылет взрослых особей из одного и того же водоема при благоприятной солнечной погоде происходит в течение нескольких дней, причем дважды в день - утром и вечером. Впрочем, часть европейских авторов считает, что отрождение имаго происходит ночью, которые совершают свой первый полет до рассвета. Расхождения во мнениях могут быть связаны как с географическим различиями, так и с банальной недостаточной изученностью процесса.
Календарно самки начинают вылетать, немного раньше самцов, и их число превосходит значительно число самцов. Превращение личинок во взрослых стрекоз происходит, как и у всех других видов этого рода, обыкновенно на растущей по берегам водоемов растительности в нескольких метрах от воды.
Окрыление происходит на растениях, растущих около берега на высоте от 10 см до 1 м, однако окончательное формирование имаго наблюдается на кустарниках и деревьях. Вылет взрослых особей из одного и того же водоема при благоприятной солнечной погоде происходит в течение нескольких дней, причем дважды в день - утром и вечером. Впрочем, часть европейских авторов считает, что отрождение имаго происходит ночью, которые совершают свой первый полет до рассвета. Расхождения во мнениях могут быть связаны как с географическим различиями, так и с банальной недостаточной изученностью процесса.
Календарно самки начинают вылетать, немного раньше самцов, и их число превосходит значительно число самцов. Превращение личинок во взрослых стрекоз происходит, как и у всех других видов этого рода, обыкновенно на растущей по берегам водоемов растительности в нескольких метрах от воды.

♀ Фото © Семён Левый, Борисовский р-н, Минская обл.
Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Brown Hawker. Aeshna grandis" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 156-157.
9. Smallshire D., Swash A. "Brown Hawker Aeshna grandis" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 112-113.
10. Iorio E. "Les habitats des espèces de la déclinaison régionale bas-normande du Plan national d’actions en faveur des Odonates : La Grande Aeschne (Aeshna grandis)". Fiche GRETIA pour la DREAL Basse-Normandie, l’Europe et l’Agence de l’Eau Seine-Normandie, 2014. 18 pp.
11. Noel N. "La Grande Aeshne en Normandie ; Aeshna grandis (Linnaeus, 1758) (Odonata, Aeshnidae)" / L’entomologiste Haut-Normand, 2013. pp. 9-13.
12. Museo Virtuale. Museo regioale di Scieze aturali della Valle d'Aosta "Aeshna grandis (Linnaeus, 1758) Brown hawker" / Bio-Montagne - Réseau d'éducation sur la biodiversité dans les zones alpines - Programma di cooperazione transfrontaliera Italia-Svizzera 2007-2013.
13. Borkenstein A., Schröter A., Jödicke R. "Matutinal mating in Aeshna grandis and A. viridis – a behavioural pair of twins prefers early-morning sex (Odonata: Aeshnidae)" / Odonatologica 46(3/4) 2017: 207-226.
14. Petzold F., Lehmann A., Lehmann G. "Braune Mosaikjungfer – Aeshna grandis (Linnaeus 1758)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 68-69.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Brown Hawker. Aeshna grandis" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 156-157.
9. Smallshire D., Swash A. "Brown Hawker Aeshna grandis" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 112-113.
10. Iorio E. "Les habitats des espèces de la déclinaison régionale bas-normande du Plan national d’actions en faveur des Odonates : La Grande Aeschne (Aeshna grandis)". Fiche GRETIA pour la DREAL Basse-Normandie, l’Europe et l’Agence de l’Eau Seine-Normandie, 2014. 18 pp.
11. Noel N. "La Grande Aeshne en Normandie ; Aeshna grandis (Linnaeus, 1758) (Odonata, Aeshnidae)" / L’entomologiste Haut-Normand, 2013. pp. 9-13.
12. Museo Virtuale. Museo regioale di Scieze aturali della Valle d'Aosta "Aeshna grandis (Linnaeus, 1758) Brown hawker" / Bio-Montagne - Réseau d'éducation sur la biodiversité dans les zones alpines - Programma di cooperazione transfrontaliera Italia-Svizzera 2007-2013.
13. Borkenstein A., Schröter A., Jödicke R. "Matutinal mating in Aeshna grandis and A. viridis – a behavioural pair of twins prefers early-morning sex (Odonata: Aeshnidae)" / Odonatologica 46(3/4) 2017: 207-226.
14. Petzold F., Lehmann A., Lehmann G. "Braune Mosaikjungfer – Aeshna grandis (Linnaeus 1758)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 68-69.
