КОРОМЫСЛО ЗЕЛЕНОЕ
Aeshna viridis Eversmann, 1836

♀ Фото © Семён Левый, Солигорский р-н, Минская обл.
Aeshna viridis Eversmann, 1836 (коромысло зеленое)
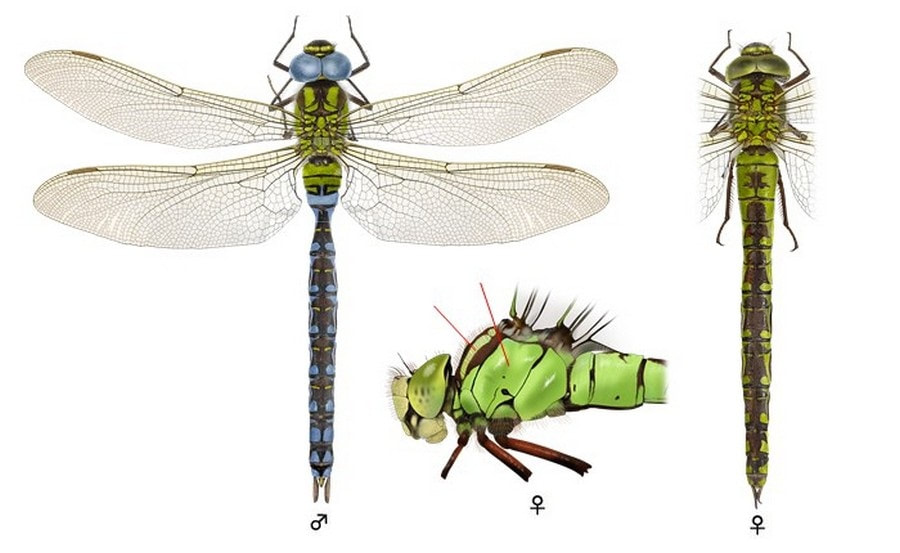
Глаза соприкасаются друг с другом на некотором отрезке. На лбу полный Т-образный черный рисунок, однако «ножка» у буквы «Т» очень тонкая. Тело пестрое. Доплечевые полосы яблочно-зеленые, полные и весьма широкие, шире, чем темные полосы между ними. Бока груди зеленые, на швах с очень узкими темными полосками. На груди сверху имеются голубые пятнышки у основания крыльев. VIII и IX тергиты брюшка сверху с парой обособленных голубых или зеленых пятен каждый. Сверху сегменты брюшка с тёмными вдавлениями в виде чёрточек и точек. Крылья прозрачные; внешняя часть с желтоватым оттенком. По мере взросления особей крылья приобретают оттенок от желтоватого до коричневатого. Костальная жилка желтая. Жилкование крыльев черное. Птеростигма темно-серая, более 3 мм. При посадке принимают вертикальную позу со свисающим вниз брюшком.
♂: Глаза светло-голубые. Брюшко также зеленое у основания, в остальном черное или очень темно-коричневое с парами больших голубых пятен и более мелкими желтыми отметинами сверху, по бокам пятна голубые. Анальный треугольник из двух ячеек. Верхние брюшные придатки имеют удлиненную вершину и не имеют суббазального зубца.
♀: Глаза зеленоватые. Рисунок такой же, как у самца, но черный цвет заменен на коричневый, все отметки зеленые и более мелкие на верхней части брюшка. Брюшко на III сегменте не сужено. Крылья с желтоватым налетом. Яйцеклад короткий, всего примерно вдвое длиннее сегмента X, задний конец его не заходит даже за конец последнего сегмента брюшка.
Неполовозрелые особи выглядят как самки.
Длина 65-75 мм, брюшко 47-54 мм, заднее крыло 38-45 мм.
Лет: конец июня - начало октября.
♂: Глаза светло-голубые. Брюшко также зеленое у основания, в остальном черное или очень темно-коричневое с парами больших голубых пятен и более мелкими желтыми отметинами сверху, по бокам пятна голубые. Анальный треугольник из двух ячеек. Верхние брюшные придатки имеют удлиненную вершину и не имеют суббазального зубца.
♀: Глаза зеленоватые. Рисунок такой же, как у самца, но черный цвет заменен на коричневый, все отметки зеленые и более мелкие на верхней части брюшка. Брюшко на III сегменте не сужено. Крылья с желтоватым налетом. Яйцеклад короткий, всего примерно вдвое длиннее сегмента X, задний конец его не заходит даже за конец последнего сегмента брюшка.
Неполовозрелые особи выглядят как самки.
Длина 65-75 мм, брюшко 47-54 мм, заднее крыло 38-45 мм.
Лет: конец июня - начало октября.

♂ Фото © Владимир Бондарь, Могилевский р-н
Населяет озера, пруды, старицы и заводи рек с плавающими гидрофитами. Тесно связан с телорезом (Stratiotes aloides), в листья которого самки обычно откладывают яйца. Однако встречается и при полном отсутствии телореза.
Преимущественно колонизирует водоемы с достаточной популяцией телореза, при условии, что они плавают на поверхности воды летом. Оптимальными являются заросли телореза, которые выступают примерно на 15-30 см над поверхностью воды и располагаются так близко друг к другу, что листья соседних растений соприкасаются или перекрываются. Это в основном озера всех размеров с мелководными участками, небольшие озера и мочажины на краю заиленных болот, торфяные выемки, старицы и заводи рек.
Стрекозы охотятся в пределах водоема и не далее 500 м от него, почти не летают в лес. Места обитания имаго включают заросли осоки и тростника, которые посещаются для кормления, спаривания и ночлега. Однако по другим сведениям, оба пола могут кормиться группами вдали от воды и отдыхать низко в грубой растительности, где их трудно заметить. Нередко залетают на территории городов. В суточной динамике наиболее активны в вечерние часы. Ночуют в кронах прибрежных деревьев и в зарослях водных макрофитов. Являются активными хищниками и в большом количестве истребляют не только комаров, мушек и мух, но и слепней. Известны случаи массового истребления ими совки-гаммы.
В эвтрофных стоячих водах регулярно наблюдается вместе с A. mixta, A. grandis, A. isoceles и чаще с Leucorrhinia pectoralis.
Преимущественно колонизирует водоемы с достаточной популяцией телореза, при условии, что они плавают на поверхности воды летом. Оптимальными являются заросли телореза, которые выступают примерно на 15-30 см над поверхностью воды и располагаются так близко друг к другу, что листья соседних растений соприкасаются или перекрываются. Это в основном озера всех размеров с мелководными участками, небольшие озера и мочажины на краю заиленных болот, торфяные выемки, старицы и заводи рек.
Стрекозы охотятся в пределах водоема и не далее 500 м от него, почти не летают в лес. Места обитания имаго включают заросли осоки и тростника, которые посещаются для кормления, спаривания и ночлега. Однако по другим сведениям, оба пола могут кормиться группами вдали от воды и отдыхать низко в грубой растительности, где их трудно заметить. Нередко залетают на территории городов. В суточной динамике наиболее активны в вечерние часы. Ночуют в кронах прибрежных деревьев и в зарослях водных макрофитов. Являются активными хищниками и в большом количестве истребляют не только комаров, мушек и мух, но и слепней. Известны случаи массового истребления ими совки-гаммы.
В эвтрофных стоячих водах регулярно наблюдается вместе с A. mixta, A. grandis, A. isoceles и чаще с Leucorrhinia pectoralis.

♀ Фото © Ольга Скробот Березовский р-н, Брестская обл.
Самцы охраняют участки вдоль берега, где доминирует телорез. Вернувшись к водоему после фазы созревания, они обустраивают там небольшие территории (максимум до 2 м) над зарослями телореза. Обычно это происходит с конца июня - начала июля. В водоемах размножения самцы отмечаются в большом количестве с позднего утра, особенно в солнечные дни. Самки прилетают к водоемам только для откладки яиц. Время лёта этого вида продолжается до конца августа, иногда до конца сентября.
Спаривание можно наблюдать вдали от водоемов в ранние послеобеденные часы, но, предположительно, оно начинается у водоема. Долгое время процесс спаривания этого вида, впрочем, как и суточная активность, оставались «белым пятном», несмотря на то, что вид относительно распространенный во всей Европе.
Считалось, что вид проявляет позднюю активность: самцы начинают свой патрульный полет поздно утром, а самки прибывают в водоем для откладывания яиц не раньше полудня. Хотя представители обоих полов встречаются в солнечные дни на плавающих зарослях телореза - единственном субстрате для откладывания яиц в Европе и на большей части ареала - спаривания здесь не зарегистрировано. Все наблюдатели сходятся во мнении относительно эффективной демонстрации отказа самки, т. е. изгибание вниз брюшка в полете. Этот сигнал откладывающих яйца самок является табу для патрулирующих самцов, которые не делают попыток сформировать пару с такими самками.
Спаривание можно наблюдать вдали от водоемов в ранние послеобеденные часы, но, предположительно, оно начинается у водоема. Долгое время процесс спаривания этого вида, впрочем, как и суточная активность, оставались «белым пятном», несмотря на то, что вид относительно распространенный во всей Европе.
Считалось, что вид проявляет позднюю активность: самцы начинают свой патрульный полет поздно утром, а самки прибывают в водоем для откладывания яиц не раньше полудня. Хотя представители обоих полов встречаются в солнечные дни на плавающих зарослях телореза - единственном субстрате для откладывания яиц в Европе и на большей части ареала - спаривания здесь не зарегистрировано. Все наблюдатели сходятся во мнении относительно эффективной демонстрации отказа самки, т. е. изгибание вниз брюшка в полете. Этот сигнал откладывающих яйца самок является табу для патрулирующих самцов, которые не делают попыток сформировать пару с такими самками.

♀ Фото © Ольга Скробот Березовский р-н, Брестская обл.
Все европейские Aeshnidae обычно начинают копуляцию в воздухе, поскольку им нужен полет для формирования «брачного кольца» сразу после создания прекопулятивного тандема. Затем совокупление продолжается на растительности в висячем положении. Однако в северо-западной Германии (Нижняя Саксония) задокументировано утреннее спаривание в характерном положении скрученного кольца. Эти кольца уникальны тем, что образуются не в полете, а сидя на растительности и в таком положении, пара не проявляет готовности убежать. Перекрученное положение, когда самец висит вниз головой, а его придатки наклонно скользят по голове самки, является результатом образования брачных колец с сидящей самкой. В это же время обнаружены раскрученные, вертикально висящие пары, которые были интерпретированы как кольца, образованные в полете, - указание на альтернативную тактику спаривания.

♂ Фото © Владимир Бондарь, Могилевский р-н
С первым утренним светом, еще до восхода солнца, оба пола прилетают к водоему размножения, пролетая над густыми зарослями телореза. Стиль их полета двух типов: хорошо известный кормовой полет и медленный, низкий, линейный и неагрессивный крейсерский полет. Совершающие крейсерский полет особи иногда образовывали брачные кольца, и пары покидали пруд. Незадолго до восхода солнца многочисленные самцы начали искать восприимчивых самок в высокой травянистой растительности у пруда. Этот неагрессивный режим полета был медленным, на высоте колена, характеризовался проникновением в густые заросли (поисковый полет). Он прекращался через 45-70 мин после восхода солнца. Предположительно, наземная растительность представляет собой основное место встреч и поисковый полет приводит к большинству спариваний. Самцы хватают отдыхающих самок и формирование кольца завершаются без дальнейшего полета, что приводит к характерному искривленному положению. Дальнейшие наблюдения подтвердили, что обнаруженные утром скрученные колеса типичны. Также имелись регистрации случайных спариваниях в период с полудня до позднего вечера. Ограничение большинства брачных игр периодом около восхода солнца опровергает миф о том, что вид имеет позднюю активность. Брачное поведение этого вида с сочетанием поискового полета, ограниченного восходом солнца, и формирования колец с отдыхающими самками кажется уникальным для стрекоз. Впрочем, несколько позже очень сходное поведение было обнаружено у A. grandis.

♀ Фото © Екатерина Морозова (Букас), Петриковский р-н, Гомельская обл.
Выделяются два основных стиля полета в утренних сумерках: кормовой и крейсерский полет. Первый не ограничивается строго прудом, а включает также кустарники вдоль береговой линии и прилегающей поляны, а второй встречается только над зарослями телореза. Оба паттерна также зарегистрированы при полетах в вечерние сумерки. Количество особей, вовлеченных в утренний пролет на водоеме, в целом, по-видимому, меньше, чем в вечерний.
Поисковый полет самцов наблюдался в период между 21 мин до и 30 мин после восхода солнца. Самцы периодически "ныряли" в растительность, а затем летали петлями, чтобы избавиться от капель воды, покрывающих крылья и тело, после проникновения в росистые заросли растений. Встречались и самцы, останавливавшиеся в период поискового полета для отдыха. Отдохнувшие самцы через некоторое время кружили, а затем отправлялись в очередной поисковый полет. Некоторые самцы остаются отдыхать на местах встреч, но как только оказывались под прямыми солнечными лучами, они разворачивались, выставляя спину под солнце.
Наземная растительность у кромки воды оказалась альтернативным и, вероятно, наиболее важным местом встреч. Самцы прилетают каждое утро около восхода солнца на все подходящие участки, характеризующиеся высокой растительностью, состоящей из Calamagrostis epigeios, Phragmites australis, Juncus effusus, Rubus sp., Urtica sp. и других. Здесь они демонстрируют поисковый полет. Наблюдения показали, что наземные места встреч не используются для ночлега. Видимо, это место выбирают восприимчивые самки, ранее участвовавшие в коллективном утреннем полете над водоемом, но не спаривавшиеся.
Поисковый полет самцов наблюдался в период между 21 мин до и 30 мин после восхода солнца. Самцы периодически "ныряли" в растительность, а затем летали петлями, чтобы избавиться от капель воды, покрывающих крылья и тело, после проникновения в росистые заросли растений. Встречались и самцы, останавливавшиеся в период поискового полета для отдыха. Отдохнувшие самцы через некоторое время кружили, а затем отправлялись в очередной поисковый полет. Некоторые самцы остаются отдыхать на местах встреч, но как только оказывались под прямыми солнечными лучами, они разворачивались, выставляя спину под солнце.
Наземная растительность у кромки воды оказалась альтернативным и, вероятно, наиболее важным местом встреч. Самцы прилетают каждое утро около восхода солнца на все подходящие участки, характеризующиеся высокой растительностью, состоящей из Calamagrostis epigeios, Phragmites australis, Juncus effusus, Rubus sp., Urtica sp. и других. Здесь они демонстрируют поисковый полет. Наблюдения показали, что наземные места встреч не используются для ночлега. Видимо, это место выбирают восприимчивые самки, ранее участвовавшие в коллективном утреннем полете над водоемом, но не спаривавшиеся.

♂ Фото © Александр Месяц, Борисовский р-н, Минская обл.
Большинство всех пар, замеченных между серединой утра и полуднем, относятся к скрученным кольцам. Это явное свидетельство регулярных спариваний в утренние сумерки, когда самцы ищут самок на наземных местах встреч. Бросается в глаза большая продолжительность положения скрученного колеса; одна пара еще висела за полчаса до полудня, что соответствует продолжительности в 7 часов и 20 минут, если колесо образовалось точно на восходе солнца. Пара не демонстрировала попыток бегства, когда наблюдатель приблизился к ней.
Нескрученные пары, обнаруженные утром, могли образоваться и на восходе солнца, но во время коллективного полета. О летающих кольцах сообщается в период между полуднем и поздним вечером. В целом, большинство спариваний происходит рано утром, когда восприимчивые самки участвуют в коллективном полете или оседают в прилегающей растительности. С другой стороны, самцы кажутся оппортунистами и спариваются всякий раз, когда обнаруживают восприимчивую самку. Об этом также свидетельствуют сообщения о кольцах, образовавшихся в месте размножения, на воде или над водой.
Нескрученные пары, обнаруженные утром, могли образоваться и на восходе солнца, но во время коллективного полета. О летающих кольцах сообщается в период между полуднем и поздним вечером. В целом, большинство спариваний происходит рано утром, когда восприимчивые самки участвуют в коллективном полете или оседают в прилегающей растительности. С другой стороны, самцы кажутся оппортунистами и спариваются всякий раз, когда обнаруживают восприимчивую самку. Об этом также свидетельствуют сообщения о кольцах, образовавшихся в месте размножения, на воде или над водой.

♀ Фото © Денис Китель.
Активность поздним утром не следует строгому графику, а зависит от погодных условий и физического состояния особей. Большинство самцов покидают место встречи с самками после утреннего поискового полета и летят к подходящим для кормления местам, как и самки, как только места их ночлега полностью прогреваются солнцем. В солнечные дни самцы начинают возвращаться в канавы примерно через 3,5 ч после восхода солнца, где совершают патрульные полеты. Самки следуют туда примерно через 2 часа для откладки яиц. Когда патрулирующие самцы встречают самок, откладывающих яйца, в основном игнорируют их. Лишь изредка наблюдаются попытки самцов приблизиться к самке, которая тут же реагирует демонстрацией отказа.
В послезакатное время оба пола встречаются в коллективном полете, но половой активности не проявляют. Сообщения о кормежке, в основном в вечернее время, не говорят о каком-либо сексуальном поведении во время охоты.
Таким образом, имеется два разных механизма объединения восприимчивых самок и самцов. Оба связаны с ранним утром. На это до сих пор не обращали внимания, и это объясняет, почему спаривание у вида в значительной степени оставалось неизвестным.
В послезакатное время оба пола встречаются в коллективном полете, но половой активности не проявляют. Сообщения о кормежке, в основном в вечернее время, не говорят о каком-либо сексуальном поведении во время охоты.
Таким образом, имеется два разных механизма объединения восприимчивых самок и самцов. Оба связаны с ранним утром. На это до сих пор не обращали внимания, и это объясняет, почему спаривание у вида в значительной степени оставалось неизвестным.
Самки откладывают яйца в листья телореза. Для этого они опускаются в воду, сползая по стеблям растений, и размещают кладки вдоль них (в основном на телорезе) на протяжении до 30 мин. Откладка яиц происходит преимущественно в жаркие полуденные часы или во второй половине дня.
Стадия яйца длится около 9 месяцев. Яйца зимуют в тканях телореза, которые осенью тонут. Примерно с сентября по апрель или май развитие яиц приостанавливается (диапауза). Предличинки отрождаются в конце апреля или начале мая.
Стадия яйца длится около 9 месяцев. Яйца зимуют в тканях телореза, которые осенью тонут. Примерно с сентября по апрель или май развитие яиц приостанавливается (диапауза). Предличинки отрождаются в конце апреля или начале мая.
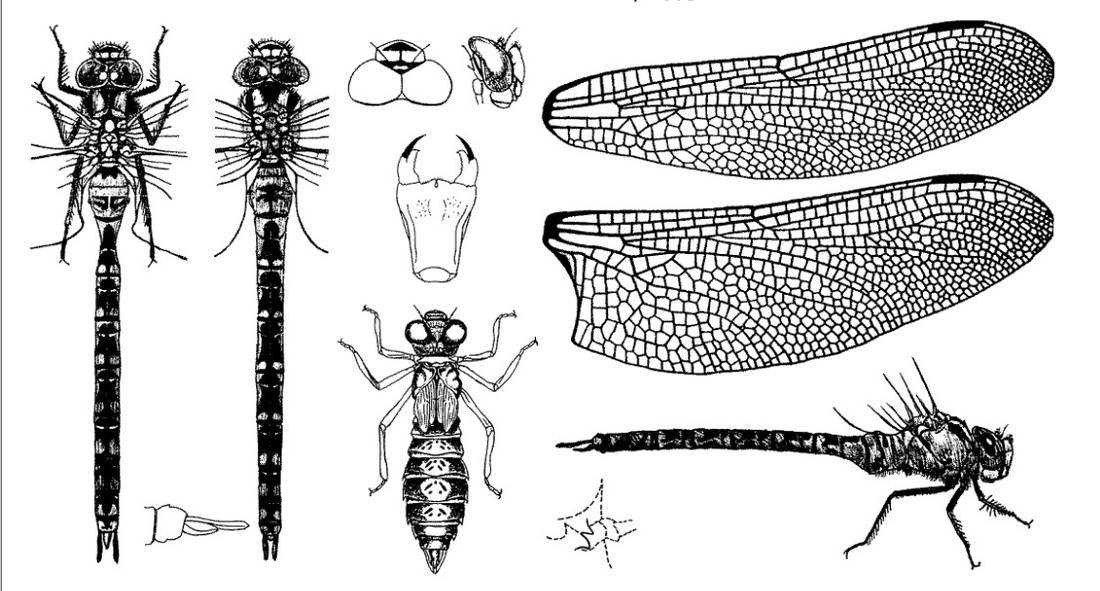
Нимфа. Темно-серая, реже коричневая, довольно неуклюжая, с относительно широким телом, напоминающая собой личинок Ае. grandis и Ае. juncea, от которых отличается формой подбородка. От личинок Ае. juncea отличается длинными четкими латеральными шипами на VI сегменте, четкой светлой полосой на голове, у личинок Ае. juncea мало заметной, и отсутствием темных срединных полос или пятен на сегментах брюшка. От личинок Ае. grandis отличается боковыми выростами переднегруди; у личинок Ае. viridis боковые выросты переднегруди почти одинаковые по величине и форме, с вершинами, направленными в одну сторону, а у Ае. grandis задние выросты шире и длиннее передних и вершины выростов направлены в разные стороны. Ноги с очень нечеткими поперечными полосами на бедрах. Брюшко относительно короткое и широкое, одноцветное, постепенно расширяющееся до VII сегмента, затем суживающееся к концу; срединные дорсальные вдавления, в виде коротких черточек и точек, темные, на IX и Х сегментах они отсутствуют. Анальная пирамида длинная; церки и дорсальный придаток такой же длины, как у личинки Ае. grandis; церки значительно длиннее IX и X сегментов, взятых вместе; церкоиды острые, цилиндрические, равны 1/2 или менее 1/2 длины церков; основная часть дорсального придатка самца коническая, с весьма острой вершиной, почти такой же длины, как церкоиды; створки яйцеклада самки не достигают заднего края IX стернита.

♀ Фото © Катерина Маковецкая, Мостовский р-н, Гродненская обл.
Лимнофилы, приурочены к водоемам с большой площадью воды (пруды, озера, карьеры, оставшиеся после разработок торфа); фитофилы, обитают среди водной растительности. Личинки ведут малоподвижный образ жизни; иногда лишь плавая или ползая. Живут между подводных стеблей телореза. Появление рыб в водоемах не исключает репродуктивного успеха A. viridis из-за укрытия личинок в пазухах листьев телореза, но часто приводит к снижению численности.
Очень прожорливы, питаются дафниями, водными осликами, личинками комаров, подёнок и др. насекомых. Набрав определённую массу, уходят в зимовку. Зимуют в пазухах листьев растения. В последнем возрасте имеют длину тела 43-44 мм. Время развития от яйца до половозрелого имаго обычно длится 2 года, при неблагоприятных условиях - 3. На последних личиночных стадиях они повторно зимуют в пазухах листьев телореза. Таким образом, в мае и начале июня на телорезе можно обнаружить одновременно два (а иногда и 3) поколения личинок.
Совершив не менее 5 линек, в июне выбираются из воды по стеблям растений, чаще рогоза и телореза и на них же превращаются в молодых стрекоз. В различных исследованиях была определена максимальная плотность отрождения до 210 экзувий/100 м². Однако плотность менее 25 экзувий/100 м² является нормой.
Наблюдаемые периоды вылета также варьировали от 48 до 63 дней. 50% всех отрождающихся стрекоз вылупились в первые 11-12 дней. Такая относительно высокая синхронизация вылупления может быть связана с определенными суммами температур или их пороговыми значениями в воде, которые должны быть достигнуты, прежде чем после 2-й зимней спячки можно будет достичь следующей личиночной стадии. Доля самцов при отрождении обычно была значительно ниже 50%.
Весной только что вылупившиеся личинки подвергаются опасности со стороны более старых личинок A. isoceles и A. grandis, а также своего собственного вида. Хищничество со стороны рыб также играет свою роль. В качестве врагов имаго рассматриваются различные виды птиц и земноводных, а также пауки.
Очень прожорливы, питаются дафниями, водными осликами, личинками комаров, подёнок и др. насекомых. Набрав определённую массу, уходят в зимовку. Зимуют в пазухах листьев растения. В последнем возрасте имеют длину тела 43-44 мм. Время развития от яйца до половозрелого имаго обычно длится 2 года, при неблагоприятных условиях - 3. На последних личиночных стадиях они повторно зимуют в пазухах листьев телореза. Таким образом, в мае и начале июня на телорезе можно обнаружить одновременно два (а иногда и 3) поколения личинок.
Совершив не менее 5 линек, в июне выбираются из воды по стеблям растений, чаще рогоза и телореза и на них же превращаются в молодых стрекоз. В различных исследованиях была определена максимальная плотность отрождения до 210 экзувий/100 м². Однако плотность менее 25 экзувий/100 м² является нормой.
Наблюдаемые периоды вылета также варьировали от 48 до 63 дней. 50% всех отрождающихся стрекоз вылупились в первые 11-12 дней. Такая относительно высокая синхронизация вылупления может быть связана с определенными суммами температур или их пороговыми значениями в воде, которые должны быть достигнуты, прежде чем после 2-й зимней спячки можно будет достичь следующей личиночной стадии. Доля самцов при отрождении обычно была значительно ниже 50%.
Весной только что вылупившиеся личинки подвергаются опасности со стороны более старых личинок A. isoceles и A. grandis, а также своего собственного вида. Хищничество со стороны рыб также играет свою роль. В качестве врагов имаго рассматриваются различные виды птиц и земноводных, а также пауки.

♀ Фото © Екатерина Морозова (Букас), Дрогичинский р-н, Брестская обл.
Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Smallshire D., Swash A. "Green Hawker. Aeshna viridis" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 144-145.
8. Bönsel A., Mauersberger R., Wachlin V. "Aeshna viridis (Eversmann, 1836) Grüne Mosaikjungfer" / FFH-Arten nach Anhang IV in Mecklenburg-Vorpommern. FFH-Code: 1048. 2010. 1-9 s.
9. Borkenstein A., Schröter A., Jödicke R. "Aeshna viridis is an early bird – matutinal matings in a crepuscular species (Odonata: Aeshnidae)" / Odonatologica 45(1/2) 2016: 37-56.
10. Borkenstein A., Schröter A., Jödicke R. "Matutinal mating in Aeshna grandis and A. viridis – a behavioural pair of twins prefers early-morning sex (Odonata: Aeshnidae)" / Odonatologica 46(3/4) 2017: 207-226.
11. Kruse A., Mauersberger R. "Grüne Mosaikjungfer – Aeshna viridis Eversmann 1836" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 78-79.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Smallshire D., Swash A. "Green Hawker. Aeshna viridis" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 144-145.
8. Bönsel A., Mauersberger R., Wachlin V. "Aeshna viridis (Eversmann, 1836) Grüne Mosaikjungfer" / FFH-Arten nach Anhang IV in Mecklenburg-Vorpommern. FFH-Code: 1048. 2010. 1-9 s.
9. Borkenstein A., Schröter A., Jödicke R. "Aeshna viridis is an early bird – matutinal matings in a crepuscular species (Odonata: Aeshnidae)" / Odonatologica 45(1/2) 2016: 37-56.
10. Borkenstein A., Schröter A., Jödicke R. "Matutinal mating in Aeshna grandis and A. viridis – a behavioural pair of twins prefers early-morning sex (Odonata: Aeshnidae)" / Odonatologica 46(3/4) 2017: 207-226.
11. Kruse A., Mauersberger R. "Grüne Mosaikjungfer – Aeshna viridis Eversmann 1836" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 78-79.
