КРАСОТКА-ДЕВУШКА
Calopteryx virgo Linnaeus, 1758

♂ Фото © Виктор Рындевич, Смолевичский р-н, Минская обл.
Calopteryx virgo Linnaeus, 1758 (красотка-девушка)
Самая крупная красотка в Европе, а также самая распространенная, заменяющая блестящую красотку в водотоках с быстрым течением. Крылья широкие, с густым жилкованием и не сужаются к черешку у основания.
♂: Тело голубовато- или зеленовато-синее, с металлическим блеском. Нижняя сторона кончика брюшка («хвостовой фонарь») темная, имеет цвет от коричневого до красноватого с обширными черными отметинами. Крылья почти целиком металлически темно-синие, блестящие, лишь вершины и основания несколько светлее (следует отметить, что у большинства особей из Беларуси эта разница обычно слабозаметна); у молодых крылья темно-бурые. Птеростигма отсутствует.
♀: Тело бронзово-зеленое, спереди блестящее, задний конец матовый. Крылья прозрачные, дымчатые, с серовато-бурыми жилками, костальная жилка металлически блестящая, зеленая. Передний край крыла с зеленым металлическим блеском. Вместо птеростигмы светлое пятно, пересеченное жилками. Оно расположено дальше от кончика крыла, чем у самок C. splendens.
Длина 45-49 мм, брюшко 34-36 мм, заднее крыло 30-31 мм.
В Европе встречаются три подвида, при этом два из них населяют южную часть континента и их появление в Беларуси маловероятно, однако чтобы было проще обращаться с европейскими справочниками, следует указать основные внешние различия. Степень окраски крыльев самцов варьируется между подвидами: ssp. virgo (северная и центральная Европа, по всей видимости, и Восточная) имеет размытые светлые участки в основании и на конце крыла, ssp. meridionalis (юго-западная) имеет только светлое основание крыльев, ssp. festiva (юго-восточная Европа) имеет крылья полностью пурпурно-синие. У самок, как правило, невозможно идентифицировать подвиды по окраске, но у некоторых крылья темнее, а рисунок похож на самца.
Период лета: (конец мая) середина июня - середина сентября.
♂: Тело голубовато- или зеленовато-синее, с металлическим блеском. Нижняя сторона кончика брюшка («хвостовой фонарь») темная, имеет цвет от коричневого до красноватого с обширными черными отметинами. Крылья почти целиком металлически темно-синие, блестящие, лишь вершины и основания несколько светлее (следует отметить, что у большинства особей из Беларуси эта разница обычно слабозаметна); у молодых крылья темно-бурые. Птеростигма отсутствует.
♀: Тело бронзово-зеленое, спереди блестящее, задний конец матовый. Крылья прозрачные, дымчатые, с серовато-бурыми жилками, костальная жилка металлически блестящая, зеленая. Передний край крыла с зеленым металлическим блеском. Вместо птеростигмы светлое пятно, пересеченное жилками. Оно расположено дальше от кончика крыла, чем у самок C. splendens.
Длина 45-49 мм, брюшко 34-36 мм, заднее крыло 30-31 мм.
В Европе встречаются три подвида, при этом два из них населяют южную часть континента и их появление в Беларуси маловероятно, однако чтобы было проще обращаться с европейскими справочниками, следует указать основные внешние различия. Степень окраски крыльев самцов варьируется между подвидами: ssp. virgo (северная и центральная Европа, по всей видимости, и Восточная) имеет размытые светлые участки в основании и на конце крыла, ssp. meridionalis (юго-западная) имеет только светлое основание крыльев, ssp. festiva (юго-восточная Европа) имеет крылья полностью пурпурно-синие. У самок, как правило, невозможно идентифицировать подвиды по окраске, но у некоторых крылья темнее, а рисунок похож на самца.
Период лета: (конец мая) середина июня - середина сентября.

♀ Фото © Сергей Одновал, Борисовский р-н, Минская обл.
Обычный вид, но встречается локально и реже, чем красотка блестящая. Стрекозы отличаются медленным порхающим полетом, напоминающим полет дневных бабочек. На распластанных крыльях не планируют. В то же время, имаго могут преодолевать большие расстояния и часто обнаруживаются вдали от водоема своего развития. В состоянии покоя держат крылья сомкнутыми, а брюшко приподнятым.
Встречаются вблизи медленно текущих ручьев и мелких речек, берега которых покрыты растительностью, предпочитая камыш и ивы. Но более крупные реки обычно избегаются, а стоячие водоемы вовсе не заселяются. Похоже, вид более толерантен к тени, чем C. splendens, однако избегает полностью затемненных водоемов (например, в еловых лесах). В целом вид привязан к природной среде с прибрежными деревьями. Кустарники около водоема служат защитой от ветра и холода. Стрекозы часто садятся на кусты и деревья, такие как ольха, растущая у кромки воды, а также на прибрежные травы на более открытых участках. Весьма осторожны и при опасности покидают места отдыха, поднимаясь высоко в кроны деревьев. Самки обычно сидят среди этой растительности, а самцы летают над поверхностью воды вдоль берега. Далеко от воды обычно не отлетают, но иногда могут отлетать и довольно далеко от места кладки яиц. При неблагоприятных погодных условиях и ночью имаго укрываются под листвой трав и деревьев.
Продолжительность жизни имаго составляет до 50 дней.
Встречаются вблизи медленно текущих ручьев и мелких речек, берега которых покрыты растительностью, предпочитая камыш и ивы. Но более крупные реки обычно избегаются, а стоячие водоемы вовсе не заселяются. Похоже, вид более толерантен к тени, чем C. splendens, однако избегает полностью затемненных водоемов (например, в еловых лесах). В целом вид привязан к природной среде с прибрежными деревьями. Кустарники около водоема служат защитой от ветра и холода. Стрекозы часто садятся на кусты и деревья, такие как ольха, растущая у кромки воды, а также на прибрежные травы на более открытых участках. Весьма осторожны и при опасности покидают места отдыха, поднимаясь высоко в кроны деревьев. Самки обычно сидят среди этой растительности, а самцы летают над поверхностью воды вдоль берега. Далеко от воды обычно не отлетают, но иногда могут отлетать и довольно далеко от места кладки яиц. При неблагоприятных погодных условиях и ночью имаго укрываются под листвой трав и деревьев.
Продолжительность жизни имаго составляет до 50 дней.

♂ Фото © Виктор Рындевич, Минский р-н
Очень интересные данные о поведении вида получены российскими специалистами во Владимирской области России вблизи г. Мстёры у р. Тара, которая имеет быстрое течение, много подводных источников и, следовательно, гораздо более холодную воду. По этой причине, в отличие от других мест, популяция красотки-девушки была изолирована от обычно сопутствующего вида C. splendens. Подробно рассмотрены различные поведенческие паттерны, виды перемещений, а также социальные отношения и процессы спаривания. Многие новые данные трактуются иначе, чем в общепринятой работе Pajunen (1966) и на которой основываются практически все современные описания западных авторов.

♀ (темная форма) Фото © Семён Левый. Борисовский р-н, Минская обл.
Расправление крыльев характерно для обоих полов. У самцов эти движения наиболее выражены, когда во время совокупления к самцу, удерживающему самку, приближается другой самец. У самок это реакция на приближение другой самки или на попытку самца совокупиться с ней. Вопреки тому, что утверждают многие источники, самец часто игнорирует это движение, а совершающая его самка не улетает и позволяет самцу схватить себя.
Расправление крыльев - это компонент позы, которую принимает сидящий самец, когда к нему приближается другой самец. Когда эта реакция достигает своего максимума, самцы блестящих красоток держат кончик брюшка в вертикальном положении, почти перпендикулярно оси тела. Самцы красоток-девушек в таких условиях лишь слегка приподнимают брюшко при расправлении крыльев. Такие же позы наблюдаются и у самок, их называют «проявлениями отказа» (полового акта).
Изгибание дистальной части брюшка. Этот компонент присутствует в упомянутом выше действии расправления крыльев у самцов. То же самое наблюдается у самцов во время танцующих полетов, являющихся стандартной частью агонистических взаимодействий. Этот элемент также наблюдался у самца, находящегося рядом с откладывающей яйца самкой.
Расправление крыльев - это компонент позы, которую принимает сидящий самец, когда к нему приближается другой самец. Когда эта реакция достигает своего максимума, самцы блестящих красоток держат кончик брюшка в вертикальном положении, почти перпендикулярно оси тела. Самцы красоток-девушек в таких условиях лишь слегка приподнимают брюшко при расправлении крыльев. Такие же позы наблюдаются и у самок, их называют «проявлениями отказа» (полового акта).
Изгибание дистальной части брюшка. Этот компонент присутствует в упомянутом выше действии расправления крыльев у самцов. То же самое наблюдается у самцов во время танцующих полетов, являющихся стандартной частью агонистических взаимодействий. Этот элемент также наблюдался у самца, находящегося рядом с откладывающей яйца самкой.

♂ Photo © Nick Uzhov / iNaturalist.org. г. Могилев. CC BY-NC 4.0
Хлопанье крыльями. В последнем случае изгибание брюшка сочетается с обычным хлопаньем крыльями. Хлопание крыльями весьма вариабельно. Скорость движения крыла в начальной фазе (расправление крыла) и максимальный угол от стандартного положения сложенных крыльев могут существенно различаться. Кроме однократного хлопания, иногда наблюдаются короткие серии из 4-5 незавершенных, отрывистых движений такого рода. У самцов C. virgo угол между крыльями перед возвращением в исходное положение обычно меньше, чем у C. splendens, так что боковые движения крыльев едва заметны, а полностью расправленные крылья отклоняются от сложенного положения менее чем на 150°.
Хлопания крыльями являются стандартными компонентами поведения самцов в трех различных контекстах: 1) отсутствие социальных партнеров; 2) перед копуляцией этот паттерн перемежается с порхающим полетом, который в таких условиях доминирует; так как самцу для подготовки к спариванию требуется максимум 1-2 минуты, то, если все идет хорошо, времени на хлопание крыльями почти не остается; 3) присутствие самца рядом с самкой во время яйцекладки; частота хлопания крыльями в этой ситуации достигает своего пика, а паузы между хлопаниями у самцов составляют в среднем 5 сек.
Эти данные подтверждают общее впечатление о более «флегматичном» поведении самцов C. virgo по сравнению с другими видами. Хлопание крыльями также выполняют самки.
Хлопания крыльями являются стандартными компонентами поведения самцов в трех различных контекстах: 1) отсутствие социальных партнеров; 2) перед копуляцией этот паттерн перемежается с порхающим полетом, который в таких условиях доминирует; так как самцу для подготовки к спариванию требуется максимум 1-2 минуты, то, если все идет хорошо, времени на хлопание крыльями почти не остается; 3) присутствие самца рядом с самкой во время яйцекладки; частота хлопания крыльями в этой ситуации достигает своего пика, а паузы между хлопаниями у самцов составляют в среднем 5 сек.
Эти данные подтверждают общее впечатление о более «флегматичном» поведении самцов C. virgo по сравнению с другими видами. Хлопание крыльями также выполняют самки.

♀ Фото © Виталий Сирота, г. Минск
Помимо стандартного линейного полета, выделяются еще 4 варианта полета, которые происходят во время социальных взаимодействий.
Танцующий полет - наиболее часто наблюдаемый. Два самца, чаще всего владельцы двух соседних участков, летают вместе примерно в 15-30 см друг от друга, иногда увеличивая расстояние до 1-2 м. Полет медленнее, чем стандартный линейный полет, порхающий и волнообразный. Ростральная часть тела приподнята, а кончик брюшка загнут. При таких взаимодействиях ни один участник не пытается приблизиться к другому ближе, чем на указанное минимальное расстояние. Нет преследователя и преследуемого, т. е. самец не домогается другого, не говоря уже о нападениях. Это взаимодействие может продолжаться в течение десятков минут и происходит над прилегающими частями соседних участков или над спорной территорией. Время от времени один из участников приземляется, но другой тут же его сгоняет, и совместный полет продолжается.
Самцы, в отличие от самцов C. splendens, часто меняют направление полета в вертикальной плоскости и выполняют действие, называемое «прыжками».
Танцующий полет - наиболее часто наблюдаемый. Два самца, чаще всего владельцы двух соседних участков, летают вместе примерно в 15-30 см друг от друга, иногда увеличивая расстояние до 1-2 м. Полет медленнее, чем стандартный линейный полет, порхающий и волнообразный. Ростральная часть тела приподнята, а кончик брюшка загнут. При таких взаимодействиях ни один участник не пытается приблизиться к другому ближе, чем на указанное минимальное расстояние. Нет преследователя и преследуемого, т. е. самец не домогается другого, не говоря уже о нападениях. Это взаимодействие может продолжаться в течение десятков минут и происходит над прилегающими частями соседних участков или над спорной территорией. Время от времени один из участников приземляется, но другой тут же его сгоняет, и совместный полет продолжается.
Самцы, в отличие от самцов C. splendens, часто меняют направление полета в вертикальной плоскости и выполняют действие, называемое «прыжками».

♂ Photo Irina Bobyleva / iNaturalist.org. Краснопольский р-н, Могилевская обл. CC BY-NC 4.0
Криволинейный полет связан с определенными взаимодействиями, которые происходят значительно реже, чем совместный танцующий полет самцов. Участники таких локомоций соблюдают дистанцию 10-15 см и летят быстро, с виражами и кривыми. Иногда они закручиваются вдоль воображаемой вертикальной оси. В отличие от порхающих полетов, которые всегда происходят непосредственно над поверхностью воды, во время криволинейного стрекозы часто одновременно поднимаются в воздух на 3-4 м. Хотя можно предположить, что конфликт начинается из-за претензий на определенную территорию, эти полеты не связаны явно с местной топографией. Если площадь радиусом ок. 2 м является начальным триггером конфликта, пути полета противников могут простираться на 10 м и более вдоль реки в обе стороны.
Эти взаимодействия очень эмоциональны. Они могут длиться 90 минут и более. Внимание обоих участников приковано к противнику. Ни один из них не реагирует на других самцов, с которыми они могут случайно столкнуться. Точно так же другие самцы не обращают внимания на пару, которая летает таким образом и несколько раз пересекает их территорию. Если к порхающему полету двух самцов могут присоединяться другие самцы, образуя таким образом временный рой из нескольких особей, то в криволинейном полете никогда не участвуют более двух самцов.
Поведение противников исключает возможность даже кратковременного перерыва в виде посадки участников, как это случается в порхающем полете. Однако, если конфликт длится час или дольше, криволинейные полеты время от времени перемежаются короткими сеансами типичного порхающего. Эти изменения внезапны и непредсказуемы.
Вопреки мнению многих авторов, российские не считают, что при кружении (следуя использованию термина западных авторов) один самец преследуется, а другой является преследователем, и им кажется, что каждый самец следует своей программе.
Самцы C. virgo переключаются с танцующего на криволинейный полет легче, чем самцы других видов. Однако в других ситуациях они кажутся более «флегматичными».
Эти взаимодействия очень эмоциональны. Они могут длиться 90 минут и более. Внимание обоих участников приковано к противнику. Ни один из них не реагирует на других самцов, с которыми они могут случайно столкнуться. Точно так же другие самцы не обращают внимания на пару, которая летает таким образом и несколько раз пересекает их территорию. Если к порхающему полету двух самцов могут присоединяться другие самцы, образуя таким образом временный рой из нескольких особей, то в криволинейном полете никогда не участвуют более двух самцов.
Поведение противников исключает возможность даже кратковременного перерыва в виде посадки участников, как это случается в порхающем полете. Однако, если конфликт длится час или дольше, криволинейные полеты время от времени перемежаются короткими сеансами типичного порхающего. Эти изменения внезапны и непредсказуемы.
Вопреки мнению многих авторов, российские не считают, что при кружении (следуя использованию термина западных авторов) один самец преследуется, а другой является преследователем, и им кажется, что каждый самец следует своей программе.
Самцы C. virgo переключаются с танцующего на криволинейный полет легче, чем самцы других видов. Однако в других ситуациях они кажутся более «флегматичными».

♂ Фото © Виталий Сирота, г. Минск
Стремительный полет очень похож по схеме на вышеупомянутые взаимодействия. Однако исполняется сольно. Это происходит в самых напряженных социальных контекстах. Например, это поведение самца, когда другой самец пытается приблизиться к самке, откладывающей яйца в пределах его участка после спаривания с хозяином территории. Такая ситуация может привести к «угону» самки вторгшимся самцом и при ней от владельца территории можно с большой долей вероятности ожидать стремительного полета. Он начинает быстро летать взад-вперед, улетая далеко за пределы своего участка. Стоит отметить, что фазы стремительного полета часто перемежаются с порхающим полетом. В ряде случаев наблюдаются быстрые полеты над водой вправо и влево, происходящие исключительно таким образом.

♂ Фото © Евгений Мисиюк / iNaturalist.org. Каменецкий р-н, Брестская обл. CC BY-NC 4.0
Порхающий полет Pajunen (1966) обозначил нейтральным термином «зависание», исходя из его кинематики. Однако далее в тексте назвал это брачным полетом. В большинстве, если не во всех последующих публикациях, использовался последний термин. Однако это вводит в заблуждение, поскольку порхающий полет наблюдается не только непосредственно перед спариванием, но и во многих других ситуациях.
Самец может выполнять порхающий полет в отсутствие самки. Нередки случаи, когда самец, вероятно, в результате общего возбуждения переходит в режим порхающего полета и движется только в этом режиме. Инерционный характер этого поведения аналогичен тому, что упоминалось для безостановочного попарного стремительного полета. Самец неоднократно улетает довольно далеко от центра своей активности и возвращается на порхающий полет. Это может продолжаться в течение 10 минут и более.
Самец может войти в это состояние, например, когда он увидел самку, которая пролетела через его территорию при неудачной попытке совокупления. Последующие множественные порхающие полеты выглядят как следствие такой внешней стимуляции. Однако переход от стандартного полета к порхающему режиму может произойти и самопроизвольно. При кратковременных повышениях общего возбуждения самец иногда конфликтует с другими, используя этот тип полета.
Порхающий полет, вне всякого сомнения, является стандартным и необходимым инструментом, обеспечивающим физический контакт между самцом и самкой во время обычного (ненасильственного) спаривания. Однако и здесь все не так однозначно. Отмечено, что иногда самец не выполнял порхающий полет и не расправлял крылья перед самкой, в отличие от данных литературных источников.
Самец может выполнять порхающий полет в отсутствие самки. Нередки случаи, когда самец, вероятно, в результате общего возбуждения переходит в режим порхающего полета и движется только в этом режиме. Инерционный характер этого поведения аналогичен тому, что упоминалось для безостановочного попарного стремительного полета. Самец неоднократно улетает довольно далеко от центра своей активности и возвращается на порхающий полет. Это может продолжаться в течение 10 минут и более.
Самец может войти в это состояние, например, когда он увидел самку, которая пролетела через его территорию при неудачной попытке совокупления. Последующие множественные порхающие полеты выглядят как следствие такой внешней стимуляции. Однако переход от стандартного полета к порхающему режиму может произойти и самопроизвольно. При кратковременных повышениях общего возбуждения самец иногда конфликтует с другими, используя этот тип полета.
Порхающий полет, вне всякого сомнения, является стандартным и необходимым инструментом, обеспечивающим физический контакт между самцом и самкой во время обычного (ненасильственного) спаривания. Однако и здесь все не так однозначно. Отмечено, что иногда самец не выполнял порхающий полет и не расправлял крылья перед самкой, в отличие от данных литературных источников.

♀ Фото © Татьяна Алешка, Воложинский р-н, Минская обл.
Амплитуда движений крыльев составляет 30-40° от горизонтальной плоскости, а частота взмахов 40-60 Гц. Крылья двигаются намного быстрее, чем при совместных полетах самцов (16 Гц). Непосредственно перед совокуплением направление полета может ненадолго измениться вперед и назад.
При готовности вступить в физический контакт с самкой, самцы занимают исходное положение, паря в 5–10 см от нее и целясь головой точно в ее сторону. Самец обычно сохраняет это положение, когда самка движется в пределах его поля зрения (часто неоднократно), следуя за ней и время от времени делая короткие движения к ней. Он не совершает таких действий, как расправление крыльев или окунание в воду. В этом смысле его поведение выглядит гораздо более жестким, чем поведение других видов.
Pajunen (1966) сообщает, что когда самцы преследуют самок, они комбинируют взмахи крыльев, как при стандартном обычном полете, и как при порхающем, а также используют промежуточные варианты. Короткие взмахи крыльев учащаются, когда самец приближается к преследуемой самке.
При готовности вступить в физический контакт с самкой, самцы занимают исходное положение, паря в 5–10 см от нее и целясь головой точно в ее сторону. Самец обычно сохраняет это положение, когда самка движется в пределах его поля зрения (часто неоднократно), следуя за ней и время от времени делая короткие движения к ней. Он не совершает таких действий, как расправление крыльев или окунание в воду. В этом смысле его поведение выглядит гораздо более жестким, чем поведение других видов.
Pajunen (1966) сообщает, что когда самцы преследуют самок, они комбинируют взмахи крыльев, как при стандартном обычном полете, и как при порхающем, а также используют промежуточные варианты. Короткие взмахи крыльев учащаются, когда самец приближается к преследуемой самке.

♂ Фото © Виталий Сирота, г. Минск
При обсуждении поведения самцов с самцами, которое обычно называют «территориальным» и «агрессивным», Панов и др. (2016) предпочитают использовать более нейтральный термин «агонистическое поведение». Два других термина кажутся менее точными. Территория самца не подходит под определение участка, принятое в литературе по поведению позвоночных и заимствованное из нее. Более того, конфликты между самцами лишены явной агрессии. Прямые контакты между самцами в виде агрессии наблюдаются лишь в редких случаях, когда самец претендует на самку, которую удерживает другой самец. В такие моменты один самец может захватить голову другого своими церками (аналогично захвату самки). Однако это происходит случайно и не выглядит как преднамеренное действие. Поэтому термины «нападение» и «борьба», широко используемые в литературе о стрекозах, кажутся неверными.

♂ Photo Irina Bobyleva / iNaturalist.org. Краснопольский р-н, Могилевская обл. CC BY-NC 4.0
Многие самцы в скоплении красоток большую часть времени проводят на своих присадах. Это может быть стебель растения, торчащий из воды, или лист, свисающий с берега. Эти отдельные присады расположены близко друг к другу: иногда ок. 1 м друг от друга, до 4-5 м. Одни присады предпочтительны, а другие занимаются редко. Максимальный срок пребывания самца на определенной присаде и ее ближайшем окружении составил 4 дня, а с однодневным перерывом - 6 дней.
Присады, которые предпочитают разные самцы, сменяющие друг друга в период размножения, можно считать центрами их активности и структурными ядрами небольших участков обитания. Присады и прилегающие к ним части нижнего яруса растительности образуют ядро участков, границы которых весьма неопределенны.
Кажется, что самцы на самом деле соревнуются за присады, а не за участки как таковые. Наблюдения позволяют предположить, что устойчивость использования самцом основной присады является достаточно хорошим предиктором его дальнейшей судьбы как владельца участка. Оказалось, что когда самец является исключительным владельцем участка, то большую часть времени он проводит на основной присаде, хотя может использовать и 2–3 второстепенных, расположенных в 1,5-2 м от основного.
Присады, которые предпочитают разные самцы, сменяющие друг друга в период размножения, можно считать центрами их активности и структурными ядрами небольших участков обитания. Присады и прилегающие к ним части нижнего яруса растительности образуют ядро участков, границы которых весьма неопределенны.
Кажется, что самцы на самом деле соревнуются за присады, а не за участки как таковые. Наблюдения позволяют предположить, что устойчивость использования самцом основной присады является достаточно хорошим предиктором его дальнейшей судьбы как владельца участка. Оказалось, что когда самец является исключительным владельцем участка, то большую часть времени он проводит на основной присаде, хотя может использовать и 2–3 второстепенных, расположенных в 1,5-2 м от основного.

♂ Photo © Stas & Lana / iNaturalist.org. г. Брест. CC BY-NC 4.0
Самец недолго остается неподвижным; он регулярно взлетает и возвращается на ту же присаду или на другую в том же районе. Различаются спонтанные взлеты и спровоцированные прилетом на участок другого самца (реже самки). Частота спонтанных взлетов следует эндогенным циклам двигательной активности, выполняя вторичную рекламную функцию. Некоторые авторы убеждены, что эти полеты всегда направлены на фуражировку, но имеются сомнения, отсюда и нейтральный термин «спонтанные взлеты».
Именно систематический характер спонтанных взлетов делает скопления самцов красоток напоминающими поведение некоторых птиц, которые сексуально привлекают самок. В пик сезона размножения, когда плотность репродуктивного скопления самцов высока, почти каждое такое действие вызывает взлет соседнего самца, затем его соседей и далее по цепной реакции. В результате образуется что-то вроде роев из 4-6 самцов. Таким образом, в скоплении постоянно поддерживается высокая динамическая плотность летающих самцов, привлекающих самок. Спонтанные взлеты, происходящие в режиме порхающего полета, являются частью прелюдии к совокуплению.
Что касается триггерных взлетов, то значительная их часть приводит к кратковременной воздушной конфронтации со вторгшимся, как правило, в совместном танцующем полете (реже, в попытке приблизиться к самке, не покидая участка). Примечательно, что когда самец тесно связан со своим участком, он избегает ненужных длительных взаимодействий с другими самцами. Об этом явно свидетельствует короткая средняя продолжительность спровоцированных взлетов, всего ок. 10 сек. При этом более чем в половине случаев (52,4%) самец после триггерного взлета возвращается на исходную присаду.
В период стабильной связи со своим участком самец часто игнорирует пролетающих мимо самцов и даже не взлетает при их прилете. Он также игнорирует конфликты соседей. В этот период действует правило «не покидать присаду».
Два владельца соседних участков регулярно взаимодействуют и участвуют в совместных танцующих полетах. Они являются спарринг-партнерами друг друга. Даже если такие взаимодействия перестают быть кратковременными и длятся несколько минут, обычно это не приводит к каким-либо изменениям в статусе владельцев участков. Вероятно, эти регулярные совместные действия могут способствовать сбросу так называемой «специфической энергии действия».
Именно систематический характер спонтанных взлетов делает скопления самцов красоток напоминающими поведение некоторых птиц, которые сексуально привлекают самок. В пик сезона размножения, когда плотность репродуктивного скопления самцов высока, почти каждое такое действие вызывает взлет соседнего самца, затем его соседей и далее по цепной реакции. В результате образуется что-то вроде роев из 4-6 самцов. Таким образом, в скоплении постоянно поддерживается высокая динамическая плотность летающих самцов, привлекающих самок. Спонтанные взлеты, происходящие в режиме порхающего полета, являются частью прелюдии к совокуплению.
Что касается триггерных взлетов, то значительная их часть приводит к кратковременной воздушной конфронтации со вторгшимся, как правило, в совместном танцующем полете (реже, в попытке приблизиться к самке, не покидая участка). Примечательно, что когда самец тесно связан со своим участком, он избегает ненужных длительных взаимодействий с другими самцами. Об этом явно свидетельствует короткая средняя продолжительность спровоцированных взлетов, всего ок. 10 сек. При этом более чем в половине случаев (52,4%) самец после триггерного взлета возвращается на исходную присаду.
В период стабильной связи со своим участком самец часто игнорирует пролетающих мимо самцов и даже не взлетает при их прилете. Он также игнорирует конфликты соседей. В этот период действует правило «не покидать присаду».
Два владельца соседних участков регулярно взаимодействуют и участвуют в совместных танцующих полетах. Они являются спарринг-партнерами друг друга. Даже если такие взаимодействия перестают быть кратковременными и длятся несколько минут, обычно это не приводит к каким-либо изменениям в статусе владельцев участков. Вероятно, эти регулярные совместные действия могут способствовать сбросу так называемой «специфической энергии действия».

Борьба за присаду самца C. virgo (сверху) с самцом C. splendens (снизу). Фото © Владимир Бондарь, Могилевский р-н
Известно, что владельцы участков часто меняются из-за того, что один самец выселяет другого. Очевидно, что между ними возникает не бой, а война нервов. Самое главное, что в этих ситуациях владелец территории не приземляется ни на основную присаду, ни на любые другие присады, даже когда они остаются свободными. Вторгшийся самец постоянно приземляется в пределах участка при любой возможности и пытается занять основную присаду.
Конфликт может быть решен в течение 60-90 минут. Если первоначальный владелец рискнет хотя бы несколько раз подряд посидеть на спорных присады, у него есть большие шансы сохранить свои права на участок. Этот шанс обратно пропорционален лётной активности такого самца. Если нарушитель каждый раз приземляется на присаду с первоначальным упорством, он побеждает в состязании и получает права на этот конкретный участок. Такие конфликты могут длиться много часов. В этом случае смена владельца является наиболее вероятным исходом. Побеждает тот, кто первым остановит нервный срыв, займет спорную площадку и благодаря этому переключится на программу «не покидать присаду».
Возвращаясь к феномену спонтанных взлетов, следует предположить, что их динамика так или иначе связана с социальным статусом самца и его текущим репродуктивным потенциалом. Замечено, что самец-претендент, выселив прежнего хозяина, сразу начинает совершать спонтанные взлеты, а количество триггерных взлетов падает, т. е. запускается автономная программа «не покидать присаду». С другой стороны, у самцов, покинувших свой участок или выселенных на периферию скопления, спонтанные взлеты значительно реже.
Конфликт может быть решен в течение 60-90 минут. Если первоначальный владелец рискнет хотя бы несколько раз подряд посидеть на спорных присады, у него есть большие шансы сохранить свои права на участок. Этот шанс обратно пропорционален лётной активности такого самца. Если нарушитель каждый раз приземляется на присаду с первоначальным упорством, он побеждает в состязании и получает права на этот конкретный участок. Такие конфликты могут длиться много часов. В этом случае смена владельца является наиболее вероятным исходом. Побеждает тот, кто первым остановит нервный срыв, займет спорную площадку и благодаря этому переключится на программу «не покидать присаду».
Возвращаясь к феномену спонтанных взлетов, следует предположить, что их динамика так или иначе связана с социальным статусом самца и его текущим репродуктивным потенциалом. Замечено, что самец-претендент, выселив прежнего хозяина, сразу начинает совершать спонтанные взлеты, а количество триггерных взлетов падает, т. е. запускается автономная программа «не покидать присаду». С другой стороны, у самцов, покинувших свой участок или выселенных на периферию скопления, спонтанные взлеты значительно реже.

Борьба за присаду самца C. virgo (сверху) с самцом C. splendens (снизу). Фото © Владимир Бондарь, Могилевский р-н
То, что при взаимодействиях типа криволинейного полета каждый участник действует автономно, и нет преследователя и преследуемого, подтверждается отсутствием четкого разграничения этого типа полета и стремительно полета, выполняемого соло. Совместные интенсивные криволинейные полеты начинаются не с уменьшения расстояния между самцами, чего следует ожидать при нападении одного самца на другого, а, наоборот, с увеличения скорости полета и увеличения длины полета. Лишь вторично стрекозы сближаются и начинают закручиваться по спирали. Точно такое же чередование увеличения расстояния (как при стремительном полете) и уменьшения расстояния между участниками наблюдается и дальше, на протяжении всего взаимодействия.
Еще одним важным моментом является то, что эти полеты не связаны с топографией участков обитания: самцы поднимаются высоко в воздух (на 3-4 м) и перемещаются постепенно, иногда на несколько десятков метров от места начала конфликта. Поэтому стремительный полет лишь является девиантной формой агрессивного поведения, бесполезной для защиты территории.
Еще одним важным моментом является то, что эти полеты не связаны с топографией участков обитания: самцы поднимаются высоко в воздух (на 3-4 м) и перемещаются постепенно, иногда на несколько десятков метров от места начала конфликта. Поэтому стремительный полет лишь является девиантной формой агрессивного поведения, бесполезной для защиты территории.

♀ Фото © Александр Анкуда, Минский р-н
При совокуплении самец обычно садится прямо на голову самки, изгибает брюшко и хватает его концом самку за шею. Затем он, активно взмахивая крыльями, поднимается вверх и приподнимает самку, так что она может коснуться половым отверстием эдеагуса самца (эта стадия занимает 6-60 сек., в среднем 23 сек.). Если попытка спаривания вынужденная, самка сопротивляется, контакт гениталий происходит очень редко. Кажется, что самки C. virgo в этом отношении более уступчивы, чем самки других видов.
Во время полового акта основание брюшка самца ритмично поднимается и опускается в исходное положение. Частота этих движений в процессе колеблется от 0,66-0,87 Гц до 0,54-0,59 Гц.
Продолжительность полового акта широко варьирует (47-165 сек., в среднем 106 сек.). Если самка восприимчива, время прекращения полового акта зависит только от самца. Продолжительность трех копуляций одного и того же меченого самца свидетельствует о снижении полового потенциала самцов со временем.
После спаривания самки некоторое время остаются на участке. Стрекоза трется концом брюшка о субстрат. У самок C. virgo промежуток времени между окончанием копуляции и началом яйцекладки значительно больше, чем у других видов (0-138 сек., в среднем 63 сек.). Предположительно, поведение половых партнеров на стадии яйцекладки у C. virgo менее связно, чем у пар C. splendens.
Во время полового акта основание брюшка самца ритмично поднимается и опускается в исходное положение. Частота этих движений в процессе колеблется от 0,66-0,87 Гц до 0,54-0,59 Гц.
Продолжительность полового акта широко варьирует (47-165 сек., в среднем 106 сек.). Если самка восприимчива, время прекращения полового акта зависит только от самца. Продолжительность трех копуляций одного и того же меченого самца свидетельствует о снижении полового потенциала самцов со временем.
После спаривания самки некоторое время остаются на участке. Стрекоза трется концом брюшка о субстрат. У самок C. virgo промежуток времени между окончанием копуляции и началом яйцекладки значительно больше, чем у других видов (0-138 сек., в среднем 63 сек.). Предположительно, поведение половых партнеров на стадии яйцекладки у C. virgo менее связно, чем у пар C. splendens.

♂ Фото © Семён Левый, Борисовский р-н, Минская обл.
После того, как самец схватил свою партнершу, пара может перелететь на другую присаду. Такие летающие тандемы гораздо чаще встречаются у C. splendens, чем у C. virgo. Летающие тандемы относятся к любой из двух категорий. Это могут быть самка и самец, которые только что совокупились и пытаются избежать конкурентов, пытающихся лишить партнерши. В других случаях в состав тандема может входить самка, поднятая самцом из воды во время яйцекладки. Летающие тандемы обычно преследуются одним или несколькими самцами и стараются покинуть зону жесткой конкуренции у поверхности воды. Тандем может пролететь довольно большое расстояние. В результате многие совокупления пар C. virgo, в том числе вынужденные, происходят в кроне деревьев на высоте до 4-5 м. Это хорошо согласуется с тем фактом, что кроны являются стандартной частью среды обитания этого вида, особенно во время ночевок.

♀ Фото © Семён Левый, Борисовский р-н, Минская обл.
Слаженное поведение партнеров после совокупления не характерно для C. virgo. Во многих случаях самец просто улетает и оставляет самку в покое. Самка, откладывающая яйца без самца, - обычное явление для этого вида. Всего за 6 часов наблюдений зафиксировано 8 таких одиночных самок, пытавшихся откладывать яйца. В 4 случаях самку схватил и унес самец, по-видимому, не тот, с которым она спаривалась до этого. Еще в двух случаях самка была унесена чужими самцами при откладке яиц в присутствии самца. Эти угоны обычно сопровождались острыми конфликтами, в которых участвовали двое из трех самцов. Однако было бы несправедливо утверждать, что самцы C. virgo никогда не проявляют поведения, напоминающего C. splendens. Но это происходит в относительно редких случаях, когда локальная плотность самцов невелика, так что владельцу участка удается монополизировать территорию вокруг самки. И даже в этих относительно редких случаях поведение самцов C. virgo отличается от закономерностей, наблюдаемых у их сородичей. Несмотря на то, что самец старается держаться рядом с самкой, его поведение лишено элементов, столь характерных для самцов C. splendens. Иногда можно наблюдать короткий порхающий полет, но трепетания крыльями никогда не бывает.

♀ Фото © Александр Анкуда, Минский р-н
Размножается в хорошо насыщенных кислородом незагрязненных ручьях и реках с умеренным или быстрым течением с илистым, гравийным или каменистым дном. Совокуплению предшествует демонстрация ухаживания - воздушный танец. Самцы привлекают самок, демонстрируя свои «хвостовые фонари» (нижняя сторона кончика брюшка) в тщательно продуманных брачных полетах. Самка откладывает яйца в одиночку в погруженные в воду ткани надводных растений и корни водных или береговых растений, окруженных водой. Также могут быть использованы для откладывания яиц плавающие тонкие корни прибрежных деревьев или древесина и ветки, лежащие в воде, а также мертвые растения.
Яйца очень мелкие, многочисленные, погружены в ткань водяных растений как торчащих над водой, так и погруженных в воду. С помощью яйцеклада самка надрезает стебель или лист водного растения и откладывает по 1 яйцу. Всего она может отложить до 300 яиц; яйца маленькие, цилиндрические, удлиненной формы, отношение длины к ширине у них 5:1. Чаще самка погружает в воду только брюшко, оставаясь над поверхностью воды. Однако есть сведения, на которые указывает и Попова (1953), что при кладке яиц самка может опускаться на дно водоема. Для Бранденбурга (Германия) указывается, что самка может пробыть под водой до 100 минут.
Яйца очень мелкие, многочисленные, погружены в ткань водяных растений как торчащих над водой, так и погруженных в воду. С помощью яйцеклада самка надрезает стебель или лист водного растения и откладывает по 1 яйцу. Всего она может отложить до 300 яиц; яйца маленькие, цилиндрические, удлиненной формы, отношение длины к ширине у них 5:1. Чаще самка погружает в воду только брюшко, оставаясь над поверхностью воды. Однако есть сведения, на которые указывает и Попова (1953), что при кладке яиц самка может опускаться на дно водоема. Для Бранденбурга (Германия) указывается, что самка может пробыть под водой до 100 минут.

♀ Фото © Владимир Бондарь. Кличевский р-н, Могилевская обл.
Нимфа. Тело удлиненное, стройное, гладкое, от желто- до красно-коричневого. Длина без антенн, включая хвостовые жабры, 30-32 мм. Голова узкая, довольно плоская, но слегка выпуклая, со слегка округлыми внешними углами; темя немного приподнятое; глазки хорошо заметные; 5 светлых отметин на лбу; глаза округлые, мало выдающиеся, умеренных размеров; антенны длинные, 6,0 мм. Маска плоская, удлиненная, в состоянии покоя простирается до основания средней пары ног. Переднегрудь квадратная, по краям темная, в центре с широкой светлой полоской, продолжающейся и на другие части груди. Крыловые чехлы длиной около 6,0 мм, на вершинах закругленные. Ноги тонкие, длинные, желтоватые; голени немного длиннее бедер; длина передних ног около 13,0 мм, средних около 18,0 мм, задних около 19,0 мм. Брюшко удлиненное, цилиндрическое, относительно широкое, незначительно суживающееся к вершине, с дорсальной стороны красно-коричневое или темно-коричневое, с вентральной - мутно красновато-коричневое, с 3 светлыми (беловатыми) дорсальными полосами: 1 центральной и 2 боковыми, проходящими по краям брюшка; на светлой центральной полосе в базальной части каждого сегмента, за исключением Х сегмента, темное узкое продольное пятно, по обе стороны которого, ближе к дистальному концу сегмента, расположено по 1 темному поперечному пятну; 2 темных менее четких пятна находятся у заднего; края каждого сегмента ближе к бокам брюшка; Х сегмент в центре приподнят в виде гребешка (киля). Хвостовые жабры длинные, значительно длиннее 1/2 длины брюшка; средняя пластинка плоская, остроконечная, длиной около 8,0 мм; 2 боковые в виде треугольников, с острыми вершинами, длиной около 9 мм; церкоиды в виде 2 маленьких отростков, расположенных между средней и боковыми жабрами; каждая из трех жабер несет по 2 темных пятна, иногда почти совсем слитых вместе.

♀ Фото © Александр Анкуда, Минский р-н
Нимфы типичные реофилы. Зимовка происходит в личиночной стадии. Развиваются в небольших мелководных реках и крупных постоянных ручьях с сильным и умеренно быстрым течением, хотя указывается и для пойменных озер и стариц. Предпочитают водоемы мелководные, богатые водной растительностью, как торчащей над водой, так и погруженной в воду. По сравнению с C. splendens, виду требуется более прохладная, равномерно прогретая вода с более высоким содержанием кислорода. Предпочитают чистые воды. Однако нимфы и взрослые насекомые регулярно встречаются в захламленной мусором и загрязненной органическими отходами реках. Поскольку личинки преодолевают небольшие расстояния в водоеме, вероятность их выживания сильно зависит от того, где именно отложены яйца. Любимые места для отдыха - подрезанные берега и тихие бухты с многочисленными убежищами. Субстрат - плавающие водные растения и свободно плавающие корни прибрежной растительности, а также зоны с опавшей листвой, принесенной течением. Личинки избегают сильных течений, малоподвижны, держатся на глубине 0,5-1 м. Могут выбираться по стеблям растений из воды и находиться на открытом воздухе до 2 дней. Причины этого явления не выяснены.
В ручьях и реках с быстрым течением часто встречается вместе с Cordulegaster boltonii и Pyrrhosoma nymphula. В других местах, где течение ослабевает, его можно увидеть с Enallagma cyathigerum, Ischnura elegans и Calopteryx splendens.
Развивается в течение 2 лет. Основной период отрождения имаго длится с начала мая до начала июня. Если погода неблагоприятная, личинки способны отложить отрождение до улучшения погоды. Вылупление начинается рано утром и продолжается до двух часов. Только что вылупившееся имаго проводит некоторое время в месте отрождения пока их покровы не затвердеют. После периода созревания около 10 дней имаго могут успешно воспроизводиться.
В ручьях и реках с быстрым течением часто встречается вместе с Cordulegaster boltonii и Pyrrhosoma nymphula. В других местах, где течение ослабевает, его можно увидеть с Enallagma cyathigerum, Ischnura elegans и Calopteryx splendens.
Развивается в течение 2 лет. Основной период отрождения имаго длится с начала мая до начала июня. Если погода неблагоприятная, личинки способны отложить отрождение до улучшения погоды. Вылупление начинается рано утром и продолжается до двух часов. Только что вылупившееся имаго проводит некоторое время в месте отрождения пока их покровы не затвердеют. После периода созревания около 10 дней имаго могут успешно воспроизводиться.

♀ Photo © natalyakaporikova / iNaturalist.org. Минский р-н. CC BY-NC 4.0

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
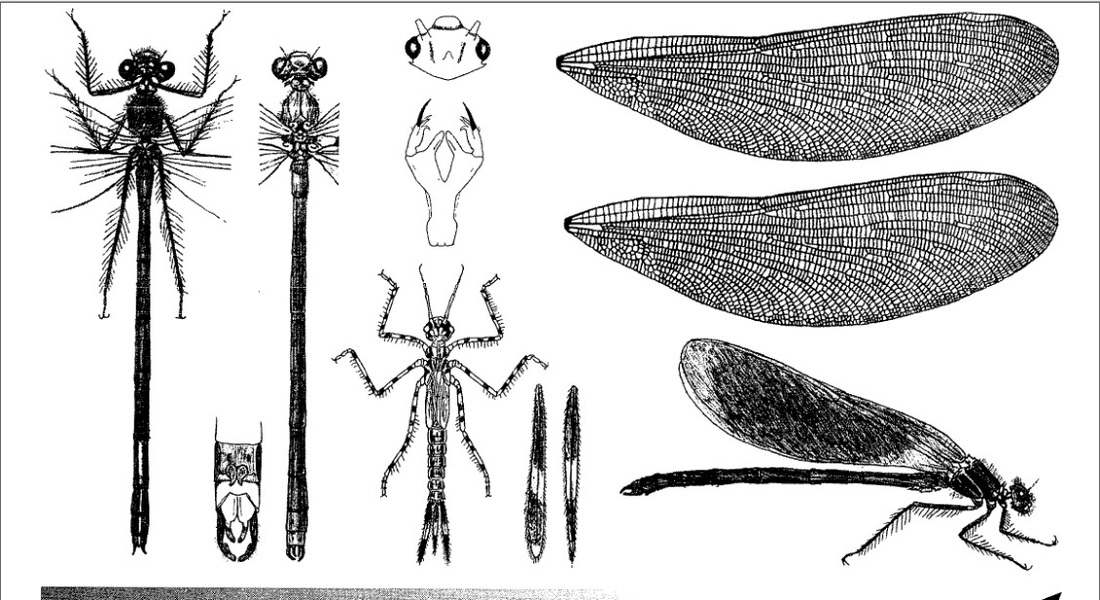
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Beautiful Demoiselle. Calopteryx virgo" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 40-41.
9. Панов Е. Н., Опаев А. С., Павлова Е. Ю., Непомнящих В. А. "Поведение двух видов равнокрылых стрекоз красотки блестящей Calopteryx splendens Harris, 1780 и красотки-девушки C. virgo Linnaeus, 1758 (Odonata, Calopterygidae)" / Russian Entomol. J. 25(1): 103-120. 2016.
10. Pajunen V. I. "Aggressive behaviour and territoriality in a population of Calopteryx virgo L. (Odonata, Calopterygidae)" / Ann. zool. fennici, 1966. Vol.3. P. 201-214.
11. Lemke M., Brauner O. "Blauflügel-Prachtlibelle – Calopteryx virgo (Linnaeus 1758)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 16-17.
12. Joest R., Wrede J. "Calopteryx virgo (Linnaeus, 1758). Blauflügel-Prachtlibelle" / im: Brockhaus et all. Atlas der Libellen Deutschlands (Odonata). Libellula Supplement 14: 18-21. 2015.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Beautiful Demoiselle. Calopteryx virgo" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 40-41.
9. Панов Е. Н., Опаев А. С., Павлова Е. Ю., Непомнящих В. А. "Поведение двух видов равнокрылых стрекоз красотки блестящей Calopteryx splendens Harris, 1780 и красотки-девушки C. virgo Linnaeus, 1758 (Odonata, Calopterygidae)" / Russian Entomol. J. 25(1): 103-120. 2016.
10. Pajunen V. I. "Aggressive behaviour and territoriality in a population of Calopteryx virgo L. (Odonata, Calopterygidae)" / Ann. zool. fennici, 1966. Vol.3. P. 201-214.
11. Lemke M., Brauner O. "Blauflügel-Prachtlibelle – Calopteryx virgo (Linnaeus 1758)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 16-17.
12. Joest R., Wrede J. "Calopteryx virgo (Linnaeus, 1758). Blauflügel-Prachtlibelle" / im: Brockhaus et all. Atlas der Libellen Deutschlands (Odonata). Libellula Supplement 14: 18-21. 2015.
