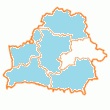
СТРЕЛКА ЗЕЛЕНОВАТАЯ или ВООРУЖЕННАЯ
Coenagrion armatum Charpentier, 1840

♂ Фото © Семён Левый, Лиозненский р-н, Витебская обл.
Coenagrion armatum Charpentier, 1840 (стрелка зеленоватая или вооруженная)
Необычно темная, стройная, нежная стрелка. Голова широкая, сверху вдвое шире своей длины. Лицо, глаза и доплечевая полоса зеленые или желтовато-зеленые. Край переднеспинки торчащий вверх и заостренный. У обоих полов характерное черное брюшко с цветными участками как у основания, так и на кончике. Черные пятна на дорсальной стороне сливающиеся. Ноги черные или темно-серые. Крылья прозрачные. Птеростигма узкая (равна 1 ячейке), светло-серая.
♂: Тело голубое, с черным рисунком. Затылочные пятна выражены хорошо, светлая полоса между ними отсутствует. Задний край переднеспинки самца округленный, посередине вытянут назад. Светлые доплечевые полосы отсутствуют, но у некоторых на их месте имеются пятна. Бока груди голубовато-зеленые. Брюшко в основном черное, голубая или сине-зеленая окраска на нем ограничена I-III и VIII-IX сегментами. Тергиты II-VII с широким черным пятном, занимающим 3/4 (и больше) членика; рисунок II тергита самца из трех пятен: среднего округлого, упирающегося в задний край тергита, и двух боковых полосок, расположенных впереди, которые, однако, могут быть слиты со средним пятном; VIII тергит голубой, IX в большей части, X сплошь черный. Нижние анальные придатки длинные щипцевидные, гораздо длиннее верхних и длиннее X членика брюшка, темные, с беловатым налетом.
♀: Затылочные пятна выражены хорошо, между ними узкая светлая полоса. Светлые доплечевые полосы узкие, но сплошные. Переднеспинка по бокам зеленая, ее задний край треугольный, посредине с узким, сильно приподнятым выступом. Светлые части тела самок могут варьироваться от светло-голубого до оливково-зеленого цвета. Неполовозрелые самки розово-оранжевые. Тергит I светлый, II с небольшим черным пятном на заднем крае, III черный с узким светлым поясом, IV-VII и IX-X сплошь черные, VIII черный в задней половине. Анальные придатки черные. Птеростигма обычно темно-серая, ее длина вдоль равна ширине или чуть длиннее.
♂: Тело голубое, с черным рисунком. Затылочные пятна выражены хорошо, светлая полоса между ними отсутствует. Задний край переднеспинки самца округленный, посередине вытянут назад. Светлые доплечевые полосы отсутствуют, но у некоторых на их месте имеются пятна. Бока груди голубовато-зеленые. Брюшко в основном черное, голубая или сине-зеленая окраска на нем ограничена I-III и VIII-IX сегментами. Тергиты II-VII с широким черным пятном, занимающим 3/4 (и больше) членика; рисунок II тергита самца из трех пятен: среднего округлого, упирающегося в задний край тергита, и двух боковых полосок, расположенных впереди, которые, однако, могут быть слиты со средним пятном; VIII тергит голубой, IX в большей части, X сплошь черный. Нижние анальные придатки длинные щипцевидные, гораздо длиннее верхних и длиннее X членика брюшка, темные, с беловатым налетом.
♀: Затылочные пятна выражены хорошо, между ними узкая светлая полоса. Светлые доплечевые полосы узкие, но сплошные. Переднеспинка по бокам зеленая, ее задний край треугольный, посредине с узким, сильно приподнятым выступом. Светлые части тела самок могут варьироваться от светло-голубого до оливково-зеленого цвета. Неполовозрелые самки розово-оранжевые. Тергит I светлый, II с небольшим черным пятном на заднем крае, III черный с узким светлым поясом, IV-VII и IX-X сплошь черные, VIII черный в задней половине. Анальные придатки черные. Птеростигма обычно темно-серая, ее длина вдоль равна ширине или чуть длиннее.

♀ Фото © Семён Левый, Шарковщинский р-н, Витебская обл.
Легко отличается от других видов рода по торчащему вверх и заостренному краю переднеспинки. Одним из отличий самца от других видов является полностью черный X сегмент брюшка. В отличие от других видов этого рода, на груди самца нет предплечевых полос, хотя могут встречаться четыре пятна. Черная маркировка на II сегменте брюшка обычно имеет боковой, иногда изолированный, отросток, идущий по обеим сторонам сегмента. Однако наиболее примечательной особенностью самцов C. armatum являются анальные придатки, у которых нижняя пара очень удлиненная. Самки большей частью тускло-черные с зеленоватыми отметинами, но они уникальны тем, что черный цвет на дорсальной поверхности II сегмента не распространяется на всю длину сегмента - передняя половина зеленоватая.
Длина 31-34 мм, брюшко 24-26 мм, заднее крыло 17-19 мм.
Лет: середина мая - конец июня.
Длина 31-34 мм, брюшко 24-26 мм, заднее крыло 17-19 мм.
Лет: середина мая - конец июня.

♂ Фото © Виктор Рындевич, окр. г. Докшицы, Витебская обл.
Вид имеет восточносибирское происхождение и распространен в Сибири (Россия), северной и восточной Европе. В Европе зарегистрирован в Великобритании, Норвегии, Швеции, Финляндии, Дании, Нидерландах, Германии, Польше, Беларуси, Украине. В Польше было известно 38 местонахождений этого вида, но только 6 из них подтверждены после 1980 г. В Украине был отмечен в 14 местах. По-видимому, всегда был редким видом, за исключением северо-западной части (Волынская область), где был более обычен. Последние регистрации на севере Украины относятся к 2000-2003 гг., однако они касались только спорадических регистраций отдельных особей, а не больших популяций. Экспедиция, посвященная повторному посещению 10 из 14 ранее известных мест весной 2007 г., не дала подтверждения ни в одном из них.
Со второй половины ХХ в. в Великобритании вид считается вымершим, хотя в начале ХХ века был обычным и встречался в больших количествах. Такая же ситуация наблюдалась и в Чехии. В Нидерландах был обнаружен в трех местах в начале ХХ века, но с 1956 г. считался вымершим, однако снова был обнаружен в 1999 г. и в настоящее время стабильные популяции присутствуют в торфяном болоте под названием «Верриббен». Аналогичная ситуация существует в Дании, где с 2005 г. были зарегистрированы две новые популяции этого вида. Следовательно, весьма вероятно, что в этих странах этот вид не исчезал, а его просто не замечали. Из-за скрытного поведения и короткого и раннего периода лета его легко не заметить во время обычных наблюдений за стрекозами.
Со второй половины ХХ в. в Великобритании вид считается вымершим, хотя в начале ХХ века был обычным и встречался в больших количествах. Такая же ситуация наблюдалась и в Чехии. В Нидерландах был обнаружен в трех местах в начале ХХ века, но с 1956 г. считался вымершим, однако снова был обнаружен в 1999 г. и в настоящее время стабильные популяции присутствуют в торфяном болоте под названием «Верриббен». Аналогичная ситуация существует в Дании, где с 2005 г. были зарегистрированы две новые популяции этого вида. Следовательно, весьма вероятно, что в этих странах этот вид не исчезал, а его просто не замечали. Из-за скрытного поведения и короткого и раннего периода лета его легко не заметить во время обычных наблюдений за стрекозами.

♀ Фото © Семён Левый, Шарковщинский р-н, Витебская обл.
Для оценки современного распространения вида в земле Шлезвиг-Гольштейн (Германия), в период с 24 апреля по 5 июня 2008 г. было проведено общенациональное обследование. Имаго C. обнаружены на 18 водоемах, расположенных на 12 участках, которые все располагались в северной части земли Шлезвиг-Гольштейн. На восьми водоемах было обнаружено более 10 имаго, тогда как на мелководном озере и в торфяной яме было зарегистрировано более 100 имаго. Вместе с тем, следует отметить, что, например, на оз. Мёвензее 06.05.2008 было насчитано более 1000 взрослых особей, а 09.05.2008 - только 180.
Тандемы обнаружены на 13 водоемах и яйцекладка в 4 из этих мест. Только одна пара была замечена в положении брачного колеса. Вид заселял мезотрофные водоемы площадью от 0,01 до 4,7 га с мелководными зонами. Большинство водоемов считались многолетними. Во всех известных случаях были водоемы были окружены болотами и вересковыми пустошами. Ситник Juncus effusus встречался, по крайней мере частично, в виде прибрежной растительности на всех занятых видом водоемах. Заросли ситника предлагали присады и использовались для укрытия и спаривания. Обломанные плавающие на поверхности воды стебли ситника даже использовались для откладки яиц.
Тандемы обнаружены на 13 водоемах и яйцекладка в 4 из этих мест. Только одна пара была замечена в положении брачного колеса. Вид заселял мезотрофные водоемы площадью от 0,01 до 4,7 га с мелководными зонами. Большинство водоемов считались многолетними. Во всех известных случаях были водоемы были окружены болотами и вересковыми пустошами. Ситник Juncus effusus встречался, по крайней мере частично, в виде прибрежной растительности на всех занятых видом водоемах. Заросли ситника предлагали присады и использовались для укрытия и спаривания. Обломанные плавающие на поверхности воды стебли ситника даже использовались для откладки яиц.
Предпочитает мелководные пруды и озера со средними по густоте зарослями хвоща (Equisetum fluviatile) или высоких и узколистных растений вроде тростника, камыша или осок, допускающих лишь ограниченное обогащение питательными веществами; иногда населяет рвы и реки с очень медленным течением. Может встречаться в заболоченных местах, даже вблизи больших озер. Общим признаком всех местообитаний является стабильный уровень грунтовых вод. Только в исключительных случаях вид встречается во временных водоемах, подверженных высыханию, из-за невозможности полного развития.
В Европе предпочтения видом местообитаний и типов водоемов различаются, иногда сильно. В общем, условная линия разделения проходит с северо-востока Европы на юго-запад, однако эта граница изменения предпочтений очень размытая. Например, в Финляндии встречается в реках и тихих ручьях, а в Польше проточные воды не заселяет. Дно водоемов также может быть разным: торфяным, песчаным или глинистым. В Финляндии встречается в разных типах водоемов, но предпочитает бассейны с глинистым дном. В Польше дно не является критической особенностью, хотя вид редко встречается в торфяных средах. В тоже время в Германии встречается почти только во временных торфяниках, хотя самая большая популяция (1000 особей) была обнаружена на небольшом озере Мювензее в песчаном грунте. Характерной чертой всех водоемов является тростниково-камышовая растительность, которой часто зарастают берега водоемов (например, Phragmites australis, Typha sp., Carex sp., Juncus sp., Equisetum limose, Cladium mariscus, Eriophorum angustifolium). Тип флоры также не имеет большого значения, но очень важна пространственная структура.
В Европе предпочтения видом местообитаний и типов водоемов различаются, иногда сильно. В общем, условная линия разделения проходит с северо-востока Европы на юго-запад, однако эта граница изменения предпочтений очень размытая. Например, в Финляндии встречается в реках и тихих ручьях, а в Польше проточные воды не заселяет. Дно водоемов также может быть разным: торфяным, песчаным или глинистым. В Финляндии встречается в разных типах водоемов, но предпочитает бассейны с глинистым дном. В Польше дно не является критической особенностью, хотя вид редко встречается в торфяных средах. В тоже время в Германии встречается почти только во временных торфяниках, хотя самая большая популяция (1000 особей) была обнаружена на небольшом озере Мювензее в песчаном грунте. Характерной чертой всех водоемов является тростниково-камышовая растительность, которой часто зарастают берега водоемов (например, Phragmites australis, Typha sp., Carex sp., Juncus sp., Equisetum limose, Cladium mariscus, Eriophorum angustifolium). Тип флоры также не имеет большого значения, но очень важна пространственная структура.

♂ Фото © Виктор Рындевич, окр. г. Докшицы, Витебская обл.
Поведение незаметное. Самцы летают низко, но мощно через относительно открытые заросли надводной растительности. Могут садиться на плавающие листья. Неоднократно наблюдалось, что тандемы и отдельные самки, отдыхающие в зарослях ситника у берега, падают при приближении наблюдателя и затем с трудом обнаруживаются в растительности.
В Шлезвиг-Гольштейне (Германия) активные взрослые особи обнаруживались между 9:30 и 19:30, т. е. в любое время дня, когда проводился контроль. Самцы постоянно держались близко к воде и предпочитали сидеть там, защищенные от ветра, на краю зарослей ситника, осоки или тростникового пояса. При небольшой плотности отдельные особи, по-видимому, использовали пространственно обособленные участки берега и неоднократно залетали на одни и те же присады. С другой стороны, большинство самок было обнаружено у внешней береговой линии в зарослях молинии или тростника. Иногда они появлялись там в большом количестве. На озере Мювензее отдельные самки улетали на расстояние до 150 м от берега на освещаемую солнцем опушку леса.
В Шлезвиг-Гольштейне (Германия) активные взрослые особи обнаруживались между 9:30 и 19:30, т. е. в любое время дня, когда проводился контроль. Самцы постоянно держались близко к воде и предпочитали сидеть там, защищенные от ветра, на краю зарослей ситника, осоки или тростникового пояса. При небольшой плотности отдельные особи, по-видимому, использовали пространственно обособленные участки берега и неоднократно залетали на одни и те же присады. С другой стороны, большинство самок было обнаружено у внешней береговой линии в зарослях молинии или тростника. Иногда они появлялись там в большом количестве. На озере Мювензее отдельные самки улетали на расстояние до 150 м от берега на освещаемую солнцем опушку леса.
Единственное наблюдавшееся в Шлезвиг-Гольштейне брачное кольцо было обнаружено в болоте Шварцбергер в 11:00 непосредственно у кромки воды в ситнике. Пара оставалась в таком положении не менее 10 минут.
Откладка яиц происходит в нижнюю сторону плавающих листьев различных макрофитов, покрывающих поверхность водоема. Самка во время этого процесса никогда не опускается вся под воду, а только опускает кончик брюшка, которым делает укол на нижней стороне плавающих листьев, куда и откладывается яйцо. Самец в большинстве случаев при кладке яиц присутствует.
В Шлезвиг-Гольштейне тандемы встречались между 12:00 и 19:30, но яйцекладка наблюдалась только между 12:00 и 15:30. Для откладывания яиц самки использовали те же участки берега, где самцы занимали присады, а иногда также появлялись с высокой плотностью.
Пары не подвергались нападению самцов своего вида. Однако взаимодействие с другими видами семейства имело место. В Хёрпфельде тандем C. armatum, пролетевший над открытой водной поверхностью, около 5 минут подвергался нападению нескольких самцов Enallagma cyathigerum, после чего пара разделилась. В Esprehmer Moor был пойман гетероспецифический тандем самца C. armatum и самки C. puella. Все наблюдаемые яйцекладки происходили в тандеме, при этом яйца откладывались либо в оторванные стебли ситника, плавающие в воде, либо на нижнюю сторону листьев живых гелофитов. 12 наблюдаемых пар откладывали яйца, используя мертвые стебли Juncus effusus, одна пара использовала мертвые стебли неустановленного вида камыша или ситника, а еще одна пара откладывала яйца на щитолистник Hydrocotyle vulgaris.
Откладка яиц происходит в нижнюю сторону плавающих листьев различных макрофитов, покрывающих поверхность водоема. Самка во время этого процесса никогда не опускается вся под воду, а только опускает кончик брюшка, которым делает укол на нижней стороне плавающих листьев, куда и откладывается яйцо. Самец в большинстве случаев при кладке яиц присутствует.
В Шлезвиг-Гольштейне тандемы встречались между 12:00 и 19:30, но яйцекладка наблюдалась только между 12:00 и 15:30. Для откладывания яиц самки использовали те же участки берега, где самцы занимали присады, а иногда также появлялись с высокой плотностью.
Пары не подвергались нападению самцов своего вида. Однако взаимодействие с другими видами семейства имело место. В Хёрпфельде тандем C. armatum, пролетевший над открытой водной поверхностью, около 5 минут подвергался нападению нескольких самцов Enallagma cyathigerum, после чего пара разделилась. В Esprehmer Moor был пойман гетероспецифический тандем самца C. armatum и самки C. puella. Все наблюдаемые яйцекладки происходили в тандеме, при этом яйца откладывались либо в оторванные стебли ситника, плавающие в воде, либо на нижнюю сторону листьев живых гелофитов. 12 наблюдаемых пар откладывали яйца, используя мертвые стебли Juncus effusus, одна пара использовала мертвые стебли неустановленного вида камыша или ситника, а еще одна пара откладывала яйца на щитолистник Hydrocotyle vulgaris.
Как уже указывалось, вид имеет восточносибирское происхождение и хотя широко распространен в лесной и лесостепной зонах Евразии, наиболее многочислен на юге Западной Сибири. Здесь при исследовании популяции стрекоз на оз. Фадиха в Барабинской лесостепи, обнаружен неизвестный ранее вариант пространственного распределения имаго после выхода из личинок. В локальной популяции на одном из озер с мощным тростниковым бордюром обнаружились две топические внутрипопуляционные группировки. Личинки одной из них развиваются в прибрежных зарослях тростника, и имаго после выплода по обычной схеме улетают на берег, личинки другой обитают среди погруженной растительности на открытом плесе, отгороженном от берега тростниковым бордюром, и имаго после окрыления не покидают плес, а проходят дополнительное питание и размножение здесь же, на месте выплода. Предположение о возможности такой несвойственной стрекозам филопатрии, выраженной в привязанности к одному плесу, возникло на основе визуальных наблюдений за поведением имаго, а также серии небольших учетов имаго и личинок.

♂ Фото © Семён Левый, Борисовский р-н, Минская обл.
Нимфа. Окраска тела коричневая, с темными точками, разбросанными по всей поверхности. Длина 22-23 мм. Голова короче, чем у личинок С. hastulatum, шириной около 4 мм; глаза серовато-зеленые, не сильно выдающиеся; антенны коричневые, более светлые по направлению к вершине; затылок с неглубокой выемкой на задней стороне. Маска в состоянии покоя простирается до основания средней пары ног; подбородок относительно короткий и широкий; передний край средней лопасти выдается посредине в виде тупого угла, стороны которого мелко зазубрены; внешний край боковых лопастей выпуклый; конечный крючок клювообразный; подвижный крючок довольно длинный, острый, искривленный. Переднегрудь дорсально с узкой желтоватой срединной продольной полосой; грудь довольно сильная, светло-коричневая, с более темными точками. Крыловые чехлы серовато-коричневые, без пятен, длиной 5,5 мм. Ноги светло-коричневые. Брюшко с многочисленными темными точками, с дорсальной стороны коричневое, с узкой желтоватой продольной срединной полосой, по краям более светлое; каждый сегмент, за исключением двух последних, с узкими желтоватыми полосами по заднему краю; брюшко с вентральной стороны светлее, чем с дорсальной. Хвостовые жаберные пластинки длинные, около 6 мм, относительно широкие, около 1,5 мм, округлые или слабо заостренные на вершине; узелковый поперечный шов не вполне прямой, по середине трахеи слабо изогнутый по направлению к основанию, и узелок на вентральном крае находится ближе к основанию, чем на дорсальном; ветвление трахей густое, но не столь древообразное, как у С. hastulatum.
Типичные фитофилы, населяют водоемы, исключительно богатые водной растительностью. Местами обитания служат разнообразные водоемы, как проточные, так и непроточные, но чаще всего это богатые водной растительностью маленькие озерки, лужицы и ямы с глинистым грунтом.
Яйца откладываются в конце весны и начале лета, а к концу осени большинство личинок завершают развитие и уходят на зимовку в финальном возрасте. Цикл развития унивольтинный.
Яйца откладываются в конце весны и начале лета, а к концу осени большинство личинок завершают развитие и уходят на зимовку в финальном возрасте. Цикл развития унивольтинный.

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
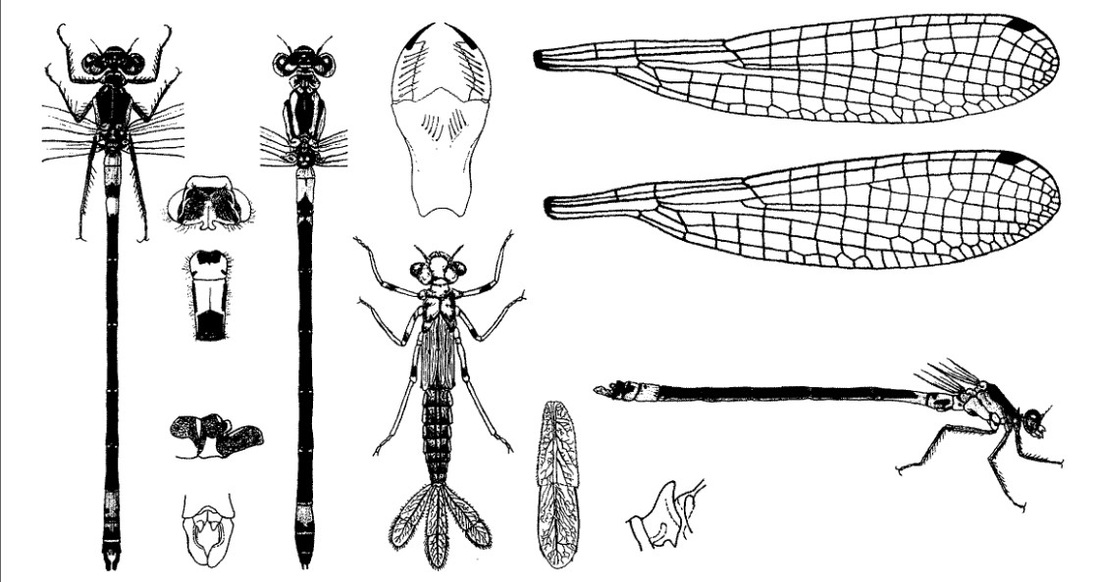
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Galliani C., Scherini R., Piglia A. "Dragonflies and Damselflies of Europe". WBA Handbooks 7, Verona, 2017: 1-352.
8. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
9. Smallshire D., Swash A. "Dark Bluet. Coenagrion armatum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 82-83.
10. Buczyński P. "On the occurrence of Coenagrion armatum (Charpentier, 1840) in Poland (Odonata: Coenagrionidae)" / Opusc. zool. flumin. 179 (2000): 1-10.
11. Khrokalo L., Krylovskaya S. "Distribution and current status of Coenagrion armatum (Charpentier, 1840) in Ukraine" / IDF-Report 13 (2008): 1-16.
12. Bouwman J., Ketelaar R. "New records of Coenagrion armatum in Schleswig-Holstein (Odonata: Coenagrionidae)" / Libellula 27 (3/4) 2008: 185-190.
13. Winkler C., Neumann H., Drews A. "Verbreitung und Ökologie von Coenagrion armatum am südwestlichen Arealrand in Schleswig-Holstein (Odonata: Coenagrionidae)" / Libellula 28 (1/2) 2009: 1-24.
14. Попова О. Н., Харитонов А. Ю. "Обнаружение топических группировок в одной из популяций стрекозы Coenagrion armatum (Charpentier, 1840)" / Сибирский экологический журнал, 2 (2014) 215–223.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Galliani C., Scherini R., Piglia A. "Dragonflies and Damselflies of Europe". WBA Handbooks 7, Verona, 2017: 1-352.
8. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
9. Smallshire D., Swash A. "Dark Bluet. Coenagrion armatum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 82-83.
10. Buczyński P. "On the occurrence of Coenagrion armatum (Charpentier, 1840) in Poland (Odonata: Coenagrionidae)" / Opusc. zool. flumin. 179 (2000): 1-10.
11. Khrokalo L., Krylovskaya S. "Distribution and current status of Coenagrion armatum (Charpentier, 1840) in Ukraine" / IDF-Report 13 (2008): 1-16.
12. Bouwman J., Ketelaar R. "New records of Coenagrion armatum in Schleswig-Holstein (Odonata: Coenagrionidae)" / Libellula 27 (3/4) 2008: 185-190.
13. Winkler C., Neumann H., Drews A. "Verbreitung und Ökologie von Coenagrion armatum am südwestlichen Arealrand in Schleswig-Holstein (Odonata: Coenagrionidae)" / Libellula 28 (1/2) 2009: 1-24.
14. Попова О. Н., Харитонов А. Ю. "Обнаружение топических группировок в одной из популяций стрекозы Coenagrion armatum (Charpentier, 1840)" / Сибирский экологический журнал, 2 (2014) 215–223.
