СТРЕЛКА КРАСИВАЯ
Coenagrion pulchellum Vander Linden, 1825

♂ Фото © Виталий Сирота, Минский р-н
Coenagrion pulchellum Vander Linden, 1825 (стрелка красивая или стрелка хорошенькая)
Голова широкая, сверху вдвое шире своей длины, сверху темная. Между затылочными пятнами имеется тонкая светлая полоса. Задний край переднеспинки с тремя примерно одинаково развитыми лопастями, хотя средняя более узкая и заостренная (особенно у самок). Лопасти обособлены хорошо. Грудь сверху темная, с двумя светлыми предплечевыми полосами, на боках груди имеется «шпора». Крылья прозрачные. Птеростигма узкая (равна 1 ячейке), одноцветная. Ноги черные или темно-серые.
♂: Основной цвет голубой. Предплечевые полосы очень узкие, обычно прерываются, образуя форму «восклицательного знака» («!»). Брюшко голубое, с тонкими черными кольцами у основания II-V сегментов. На II тергите брюшка толстое черное пятно, которое упирается в задний край тергита, образует обращенную вперед развилку (как винный бокал в поперечном сечении) и доходит своими боковыми ветвями почти до переднего края членика; иногда по середине развилки имеется еще средняя черная полоска; на III-V сегментах дорсально на базальной части заостренные черные пятна, от которых латерально отходит черная линия, проходящая вдоль сегмента; сегменты VI и IX почти черные, VIII - голубой, VII и X - полностью черные. Расширение черных пятен III-VI сегментов брюшка может варьировать (1/4 сегмента, 1/2 и даже больше). Верхние анальные придатки самцов соприкасаются в базальной части и не имеют апикального зубца; дорсальный выступ нижних придатков очень тонкий.
♀: Голубовато- или желтовато-зеленая с черным рисунком, выраженным интенсивнее, чем у самца. Самки в двух формах: у одной черные пятна тергитов II-VII не доходят до переднего края членика, а занимают 2/3 или 3/4 его; у другой пятна тергитов II-X пересекают весь членик от переднего до заднего края. У некоторых темных особей может быть преимущественно голубой сегмент VIII (напоминая C. armatum).
♂: Основной цвет голубой. Предплечевые полосы очень узкие, обычно прерываются, образуя форму «восклицательного знака» («!»). Брюшко голубое, с тонкими черными кольцами у основания II-V сегментов. На II тергите брюшка толстое черное пятно, которое упирается в задний край тергита, образует обращенную вперед развилку (как винный бокал в поперечном сечении) и доходит своими боковыми ветвями почти до переднего края членика; иногда по середине развилки имеется еще средняя черная полоска; на III-V сегментах дорсально на базальной части заостренные черные пятна, от которых латерально отходит черная линия, проходящая вдоль сегмента; сегменты VI и IX почти черные, VIII - голубой, VII и X - полностью черные. Расширение черных пятен III-VI сегментов брюшка может варьировать (1/4 сегмента, 1/2 и даже больше). Верхние анальные придатки самцов соприкасаются в базальной части и не имеют апикального зубца; дорсальный выступ нижних придатков очень тонкий.
♀: Голубовато- или желтовато-зеленая с черным рисунком, выраженным интенсивнее, чем у самца. Самки в двух формах: у одной черные пятна тергитов II-VII не доходят до переднего края членика, а занимают 2/3 или 3/4 его; у другой пятна тергитов II-X пересекают весь членик от переднего до заднего края. У некоторых темных особей может быть преимущественно голубой сегмент VIII (напоминая C. armatum).

© Владимир Бондарь, Могилевский р-н
Идентификация, как и большинства других видов этого рода, требует тщательного изучения. Черная отметка на II сегменте брюшка обычно имеет ножку и обычно намного толще, чем у самцов C. puella, на которых похож этот вид. Иногда, однако, эта ножка отсутствует. Черная отметка на дорсальной поверхности IX сегмента почти всегда более обширна, чем у C. puella. Форма анальных придатков имеет диагностическое значение. Различают две формы самок: одна темная с зеленовато-желтыми отметинами по бокам груди и брюшка, а другая - голубая, у которой черные отметины на дорсальной поверхности брюшка более обширны, чем у самца, и черная отметка на II сегменте брюшка внешне напоминает форму листа чертополоха. Эта форма почти идентична голубой форме самки C. puella, но может быть отделена от нее трехлопастным задним краем переднеспинки.
Длина 34-38 мм, брюшко 25-30 мм, заднее крыло 16-21 мм.
Лет: май - начало августа (сентябрь). Можно предположить, что вид имеет в году 2 поколения.
Длина 34-38 мм, брюшко 25-30 мм, заднее крыло 16-21 мм.
Лет: май - начало августа (сентябрь). Можно предположить, что вид имеет в году 2 поколения.

♂ Фото © Виталий Сирота, г. Минск
Населяет все типы стоячих и слабо проточных водоемов с богатой водной растительностью и не слишком кислой водой. Места часто представляют собой защищенные, но не затененные небольшие мезотропные водоемы с окружающими лесными массивами. Участки на востоке Великобритании часто напоминают болотистую местность. Описаны места размножения, такие как заросшие пастбищные канавы, болота, пруды и озера, где вид часто встречается вместе с Brachytron pratense и Sympetrum sanguineum. В Ирландии предпочитает низинные болота, небольшие озера и пруды; наиболее распространен в мезотропных местах, реже в эвтрофных. Вероятно, больше зависит от надводной растительности, чем C. puella.
Для Германии также указывается очень широкий диапазон типов водоемов: озера, пруды, торфяные болота, старые карьеры, участки рек с медленным течением, каналы, широкие канавы и старицы рек. Поскольку вид очень хорошо приспособлен к сосуществованию с рыбами, он гораздо чаще, чем C. puella заселяет более крупные акватории. Прямых свидетельств размножения во временных водоемах нет.
Большинство аспектов поведения похожи на C. puella, взрослых особей часто можно найти в пышной растительности недалеко от кромки воды. В ветреную погоду часто обнаруживаются укрывающимися в кустах с подветренной стороны. По-видимому, вид не является территориальным и может встречаться при высокой плотности. Копуляция занимает 10-15 минут. После совокупления на берегу пары остаются в тандеме. Яйцекладка обычно начинается в полдень.
Для Германии также указывается очень широкий диапазон типов водоемов: озера, пруды, торфяные болота, старые карьеры, участки рек с медленным течением, каналы, широкие канавы и старицы рек. Поскольку вид очень хорошо приспособлен к сосуществованию с рыбами, он гораздо чаще, чем C. puella заселяет более крупные акватории. Прямых свидетельств размножения во временных водоемах нет.
Большинство аспектов поведения похожи на C. puella, взрослых особей часто можно найти в пышной растительности недалеко от кромки воды. В ветреную погоду часто обнаруживаются укрывающимися в кустах с подветренной стороны. По-видимому, вид не является территориальным и может встречаться при высокой плотности. Копуляция занимает 10-15 минут. После совокупления на берегу пары остаются в тандеме. Яйцекладка обычно начинается в полдень.

♂ Фото © Олег Алексевич, г. Брест
Яйца откладываются главным образом на нижнюю сторону плавающих и погруженных в воду листьев водных растений, а также в ткани отживших растений и в бревна сплавного леса. При кладке яиц самец всегда сопровождает самку, которая, как правило, под воду не спускается, а только погружает брюшко, чтобы яйцекладом пробуравливать дырочки для яиц на нижней стороне листьев. Яйца обычно откладываются концентрическими кругами (чаще всего в 3 круга) на равномерных расстояниях друг от друга, кончики их всегда выступают под эпидермой листьев. Отмечено откладывание яиц в листья кувшинок, пробуравленных во многих местах жуком Donaciа crassipes. Самка, вероятно, использует эти отверстия для погружения брюшка в воду, и, кроме того, они являются центром, из которого она наносит по радиусу правильные концентрические круги, водя брюшком наподобие циркуля.

♂ Фото © Владимир Бондарь, г. Могилев
Как и у C. puella, у вида нередко встречаются большие скопления яйцекладущих пар. Тандемы, откладывающие яйца, объединяются, потому что привлекают друг друга, чтобы защитить себя от одиночных самцов, которые нападают на тандемы и пытаются отделить самку. Кроме того, совместное пребывание в группе также снижает риск стать жертвой хищников. Рыба и лягушка не способны поймать более одного тандема за раз. Нападение на один тандем отпугивает остальных. Тот факт, что некоторые сородичи уже взлетают, может насторожить группу.
Тандем пролетает над поверхностью воды и приземляется на плавающий объект. Сразу же после того, как самка изогнула брюшко, происходит яйцекладка или тестирование субстракта. Если самка не изогнула брюшко сразу после приземления, она вообще не изогнет его во время этой посадки. Если пара потревожена, самец вытаскивает самку из воды, у последней часто бывает еще искривленное брюшко.
Отдохнув, опробовав субстрат или отложив яйца, тандемы взлетают, после чего неоднократно садятся на другие участки. Что касается больших плавающих листьев кувшинки и кубышки, то тандемы располагаются на их краю, кроме случаев, если эти листью уже пробуравлены жуком Donaciа crassipes. В результате число тандемов, встречавшихся на одном листе кубышки желтой, может достигать 8. Но не все они откладывают яйца. Некоторые самки из тандемов держат брюшко прямо. В течение дня групповые яйцекладки перемещаются с одного участка на другой.
Тандем пролетает над поверхностью воды и приземляется на плавающий объект. Сразу же после того, как самка изогнула брюшко, происходит яйцекладка или тестирование субстракта. Если самка не изогнула брюшко сразу после приземления, она вообще не изогнет его во время этой посадки. Если пара потревожена, самец вытаскивает самку из воды, у последней часто бывает еще искривленное брюшко.
Отдохнув, опробовав субстрат или отложив яйца, тандемы взлетают, после чего неоднократно садятся на другие участки. Что касается больших плавающих листьев кувшинки и кубышки, то тандемы располагаются на их краю, кроме случаев, если эти листью уже пробуравлены жуком Donaciа crassipes. В результате число тандемов, встречавшихся на одном листе кубышки желтой, может достигать 8. Но не все они откладывают яйца. Некоторые самки из тандемов держат брюшко прямо. В течение дня групповые яйцекладки перемещаются с одного участка на другой.

♀♀ Фото © Владимир Бондарь, Могилевский р-н
Если одиночный самец приближается к тандему, откладывающему яйца, самец из пары защищаясь вибрирует крыльями. Одиночки лишь ненадолго приземляются на грудь тандемного самца и никогда напрямую не нападают на самку.
Рыбы, ищущие пищу под листьями или плавающие чуть ниже поверхности, часто нарушают откладывание яиц тандемами. Когда группы яйцекладущих тандемов потревожены, некоторые реагируют не так быстро, как другие. Если брюшко самки все еще застревало на нижней стороне листа, то тандему часто удавалось взлететь только после большого усилия. Иногда тандем разъединяется после того, как его потревожили. Например, описывается как самка застряла между двумя листьями. Самец попытался подбросить ее в воздух. После того, как ему это не удалось, он отделился от нее и улетел.
Взлетающие тандемы часто преследуются одиночными самцами и интенсивно атакуются.
Рыбы, ищущие пищу под листьями или плавающие чуть ниже поверхности, часто нарушают откладывание яиц тандемами. Когда группы яйцекладущих тандемов потревожены, некоторые реагируют не так быстро, как другие. Если брюшко самки все еще застревало на нижней стороне листа, то тандему часто удавалось взлететь только после большого усилия. Иногда тандем разъединяется после того, как его потревожили. Например, описывается как самка застряла между двумя листьями. Самец попытался подбросить ее в воздух. После того, как ему это не удалось, он отделился от нее и улетел.
Взлетающие тандемы часто преследуются одиночными самцами и интенсивно атакуются.

♀ Фото © Семён Левый, Березовский р-н, Брестская обл.
Нимфа. Окраска тела зелено-желтоватая или желто-коричневая, с множеством темных точек. Личинки полупрозрачные, благодаря чему становятся почти незаметными среди водной растительности. Тело гладкое, стройное, длиной 19-21 мм, шириной 3-3,5 мм. Голова в 2 раза шире своей длины, округлая впереди, со светлым округлым пятном на лбу, с 2 светлыми хорошо заметными позади глаз суживающимися постантеннальными бороздками: глаза выпуклые, помещаются в передних боковых углах головы; затылок относительно узкий, задние углы округлые, не выдаются, покрытые шипиками; внешние стороны затылка скошены; задний край вогнутый; вся поверхность затылка усеяна многочисленными темными точками. Подбородок маски длиной около 2,5 мм, шириной у вершины около 1,75 мм, приблизительно за серединой резко суживается к основанию: передний край средней лопасти выдается в центре в виде тупого угла и тонко зазубрен. Переднегрудь небольшая, около 2 мм шириной, с тупоугольными темными сторонами, узкая впереди, выпуклая сзади, покрытая множеством темных точек. Крыловые чехлы короткие, 5 мм. Ноги тонкие, светлые; на бедрах в дистальной части имеется не особенно ясное темное кольцо; лапки и места сочленений также темные; длина передних ног около 6 мм, средних около 6,5 мм, задних около 9,25 мм. Брюшко цилиндрическое, постепенно суживающееся к концу, со светлой продольной срединной дорсальной полосой; боковые края также светлые; вся остальная поверхность брюшка покрыта множеством темных точек; межсегментальные пространства с чередующимися темными и светлыми пятнами; Х сегмент обрамлен шипиками, дистальный край его «выемкой; с вентральной стороны брюшко светлее, чем с дорсальной. Хвостовые жаберные пластинки ланцетовидные, округлые на вершине и с очень маленьким тупым острием, длиной около 5 мм, шириной около 2 мм; узелки довольно четкие; поперечный узелковый шов изогнутый и косой, делит пластинку на 2 части: более узкую, базальную, внешние края которой вооружены шипиками, и более широкую дистальную, обрамленную волосками; главный трахейный ствол ясный, обыкновенно проходит не по середине пластинки, а ближе к дорсальному краю; разветвление трахей четкие; пластинки желтовато-коричневые, с более темным поперечным пятном в области поперечного узелкового шва. Створки яйцеклада самки доходят почти до дистального края Х тергита.

♂ Фото © Семён Левый, г. Могилев
Фаза яйца у С. pulchellum, как и у большинства видов этого рода, короткая. В искусственных условиях из свежеотложенных яиц через 2-3 недели вылупляются личинки.
Личинки обитают в самых разнообразных водоемах: речных заводях, старицах, озерах, прудах, карьерах, осоковых и торфяных болотах, лужах, но предпочитают все же стоячие водоемы с глинистым дном и с богатой водной растительностью, в которой они держатся на глубине 0,1-0,5 м. Как правило, водоемы имеют общую более зрелую стадию сукцессии, так что органические отложения встречаются в местообитаниях вплоть до береговой зоны, тростникового пояса, плавающего листового пояса и прибрежных лесов. Поскольку вид очень хорошо приспособлен к сосуществованию с рыбами, он гораздо чаще, чем C. puella заселяет более крупные акватории.
Зимует. Цикл развития (фазы яйца и личинки) вида продолжается около 1 года.
Имаго обычно начинают отрождаться в первую неделю мая или через несколько дней после долгих холодных зим. Через несколько дней после начала вылета в оптимальных местообитаниях часто происходит массовое отрождение, а первую кладку яиц иногда можно наблюдать уже в начале мая. Отрождение обычно заканчивается в середине июня; последние свидетельства появления неокрашенных имаго относятся к началу июля. С конца июня или начала июля численность имаго у водоемов резко снижается.
Личинки обитают в самых разнообразных водоемах: речных заводях, старицах, озерах, прудах, карьерах, осоковых и торфяных болотах, лужах, но предпочитают все же стоячие водоемы с глинистым дном и с богатой водной растительностью, в которой они держатся на глубине 0,1-0,5 м. Как правило, водоемы имеют общую более зрелую стадию сукцессии, так что органические отложения встречаются в местообитаниях вплоть до береговой зоны, тростникового пояса, плавающего листового пояса и прибрежных лесов. Поскольку вид очень хорошо приспособлен к сосуществованию с рыбами, он гораздо чаще, чем C. puella заселяет более крупные акватории.
Зимует. Цикл развития (фазы яйца и личинки) вида продолжается около 1 года.
Имаго обычно начинают отрождаться в первую неделю мая или через несколько дней после долгих холодных зим. Через несколько дней после начала вылета в оптимальных местообитаниях часто происходит массовое отрождение, а первую кладку яиц иногда можно наблюдать уже в начале мая. Отрождение обычно заканчивается в середине июня; последние свидетельства появления неокрашенных имаго относятся к началу июля. С конца июня или начала июля численность имаго у водоемов резко снижается.

© Владимир Бондарь, Быховский р-н, Могилевская обл.

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
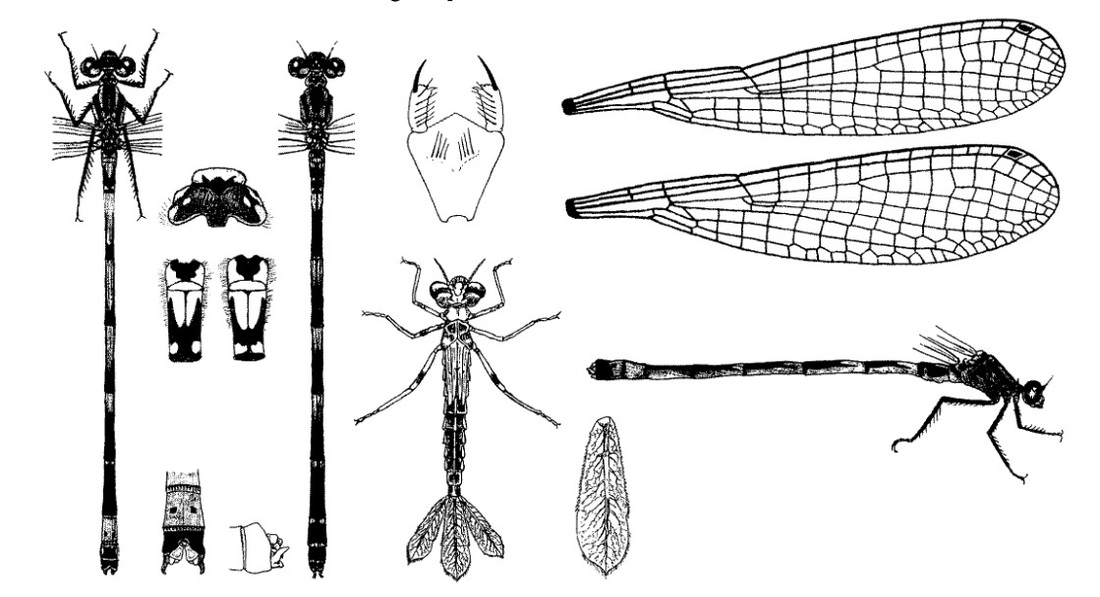
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Variable Bluet. Coenagrion pulchellum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 72-73.
9. Smallshire D., Swash A. "Variable Damselfly. Coenagrion pulchellum" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 88-89.
10. Martens A. "Aggregation of tandems in Coenagrion pulchellum (Van der Linden, 1825) during oviposition (Odonata : Coenagrionidae)" / Zool. Anz. 223 (1989) 1/2, S. 121-128.
11. Martín R. "Coenagrion pulchellum (Van der Linden, 1825) (Insecta, Odonata, Coenagrionidae) en la Península Ibérica" / Boletín de la Sociedad Entomológica Aragonesa (S.E.A.), nº 48 (30/06/2011): 493-495.
12. Tyrrell M. "Coenagrion pulchellum (Vander Linden) (Variable Damselfly in Northamptonshire VC32: A review of preferred breeding habitats" / Journal of the British Dragonfly Society, Volume 35 No. 2, 2019. pp. 70-78.
13. Mauersberger R. "Fledermaus-Azurjungfer – Coenagrion pulchellum (Vander Linden 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 46-47.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Variable Bluet. Coenagrion pulchellum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 72-73.
9. Smallshire D., Swash A. "Variable Damselfly. Coenagrion pulchellum" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 88-89.
10. Martens A. "Aggregation of tandems in Coenagrion pulchellum (Van der Linden, 1825) during oviposition (Odonata : Coenagrionidae)" / Zool. Anz. 223 (1989) 1/2, S. 121-128.
11. Martín R. "Coenagrion pulchellum (Van der Linden, 1825) (Insecta, Odonata, Coenagrionidae) en la Península Ibérica" / Boletín de la Sociedad Entomológica Aragonesa (S.E.A.), nº 48 (30/06/2011): 493-495.
12. Tyrrell M. "Coenagrion pulchellum (Vander Linden) (Variable Damselfly in Northamptonshire VC32: A review of preferred breeding habitats" / Journal of the British Dragonfly Society, Volume 35 No. 2, 2019. pp. 70-78.
13. Mauersberger R. "Fledermaus-Azurjungfer – Coenagrion pulchellum (Vander Linden 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 46-47.
