СТРЕЛКА ИЗЯЩНАЯ
Ischnura elegans Vander Linden, 1820

♂ Фото © Виталий Сирота, Минский р-н
Ischnura elegans Vander Linden, 1820 (стрелка изящная)
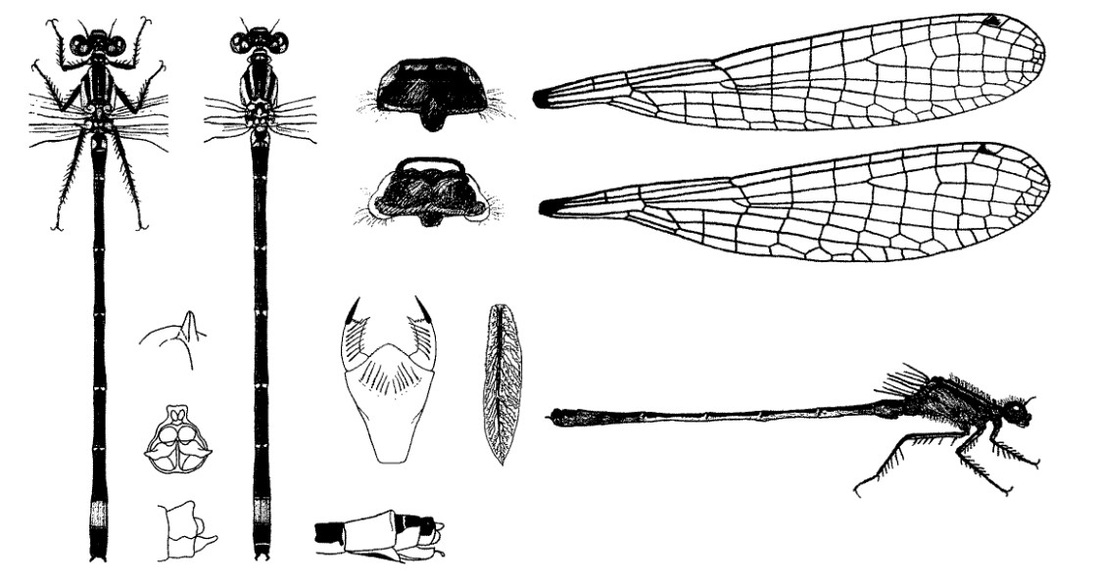
Голова широкая, сверху вдвое шире своей длины, черная. Лоб округлен. На затылке два светлых округленных пятна. Задний край переднеспинки сильно приподнят посередине и торчит в виде гребешка, однако у некоторых самок этот «шип» иногда может отсутствовать. Брюшко дорсально бронзово-черное, кроме светлого сегмента VIII. Бока сегментов I, II, низ VII-X - также светлые. Птеростигма на всех крыльях почти одинаковой величины, узкая (равна 1 ячейке), но более чем в 2 раза длиннее ширины. Ноги черные или темно-серые.
♂: Глаза, голова и грудь сверху черная. Светлый цвет голубой, включая узкие предплечевые полосы, брюшной сегмент VIII и остальные светлые части. Брюшко черное, с узкими перевязями, на конце с большим голубым "фонарем". Сильно приподнят задний край X членика брюшка. Крылья прозрачные. Птеростигма двухцветная: в основании черная, к вершине - светлая. При осмотре сзади кончики внутренних ветвей верхних придатков параллельны.
♀: Черный рисунок такой же, как у самца. Свойственен полиморфизм (см. ниже) и имеется три формы окраски в зрелом возрасте. На вентральной стороне VIII сегмента имеется острый шип. Анальные придатки черные. Птеростигма более или менее двухцветная, по крайней мере, частично светлая; имеет менее четкий рисунок, чем у самца.
Длина 30-34 мм, брюшко 24-29 мм, заднее крыло 16-18 мм.
Лет: конец мая - середина сентября; массовый лёт происходит, вероятно, с июня до последней декады августа. Вид имеет весеннее и осеннее поколения.
♂: Глаза, голова и грудь сверху черная. Светлый цвет голубой, включая узкие предплечевые полосы, брюшной сегмент VIII и остальные светлые части. Брюшко черное, с узкими перевязями, на конце с большим голубым "фонарем". Сильно приподнят задний край X членика брюшка. Крылья прозрачные. Птеростигма двухцветная: в основании черная, к вершине - светлая. При осмотре сзади кончики внутренних ветвей верхних придатков параллельны.
♀: Черный рисунок такой же, как у самца. Свойственен полиморфизм (см. ниже) и имеется три формы окраски в зрелом возрасте. На вентральной стороне VIII сегмента имеется острый шип. Анальные придатки черные. Птеростигма более или менее двухцветная, по крайней мере, частично светлая; имеет менее четкий рисунок, чем у самца.
Длина 30-34 мм, брюшко 24-29 мм, заднее крыло 16-18 мм.
Лет: конец мая - середина сентября; массовый лёт происходит, вероятно, с июня до последней декады августа. Вид имеет весеннее и осеннее поколения.

♀ Фото © Виталий Сирота, Минский р-н
Различные цветовые формы имеют и самцы, однако у них формы возрастные. Самцы меняют цвет с зеленоватого (ювенильные), на бирюзовый (зрелые) и голубой (старые). Часто указывают только первую и последнюю формы. В лабораторных условиях первая смена окраски происходит через 14 дней, а вторая – через 20 дней. По данным о повторной поимки получены значения 7 дней и 20 дней для обоих периодов. Очевидно, что первый период недооценен методом отлова-повторного отлова, так как неполовозрелых особей у воды мало, а тенеральные отмечаются редко.
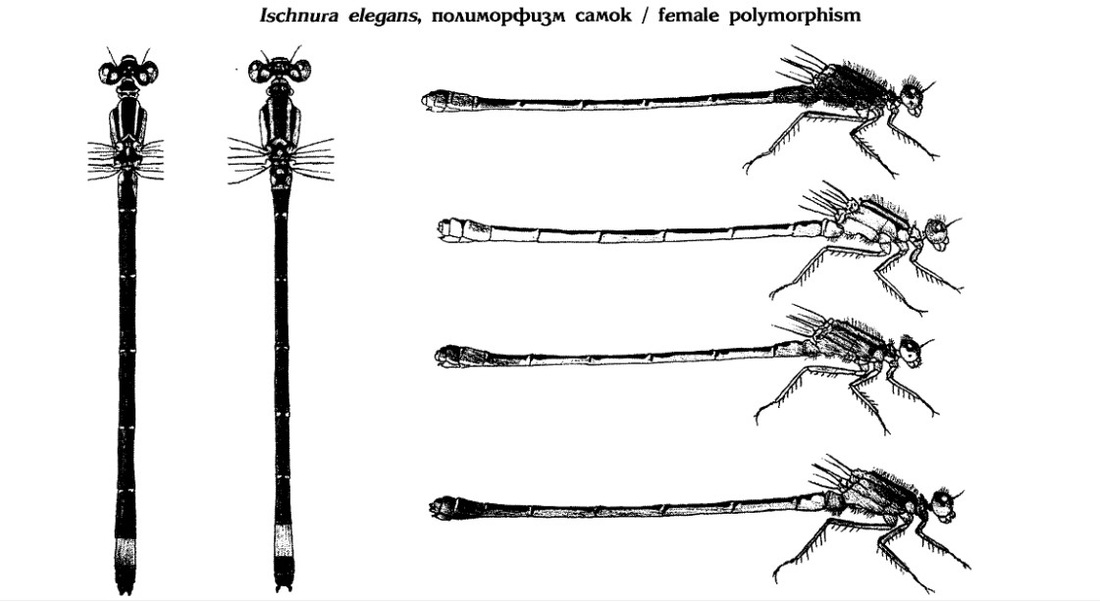
Изменения окраски самок и полиморфизм сложнее. Они имеют 5 форм, 2 у неполовозрелых (violacea и rufescens) и 3 у зрелых. Неполовозрелые с фиолетовыми боками груди (violacea) в зрелом возрасте приобретают либо окраску самцов (андроморфную или typica), либо оливково-зеленую грудь и коричневый сегмент VIII (infuscans). Другая неполовозрелая форма (rufescens) имеет оранжево-розовые бока груди, которые по мере созревания становятся светло-коричневыми с тускло-коричневым сегментом VIII (rufescens-obsoleta, ранее называвшуюся infuscans-obsoleta): обе эти формы имеют нечеткие темные линии под широкими светлыми предплечевыми полосами или вовсе их не имеют.
Изменения окраски самок и полиморфизм сложнее. Они имеют 5 форм, 2 у неполовозрелых (violacea и rufescens) и 3 у зрелых. Неполовозрелые с фиолетовыми боками груди (violacea) в зрелом возрасте приобретают либо окраску самцов (андроморфную или typica), либо оливково-зеленую грудь и коричневый сегмент VIII (infuscans). Другая неполовозрелая форма (rufescens) имеет оранжево-розовые бока груди, которые по мере созревания становятся светло-коричневыми с тускло-коричневым сегментом VIII (rufescens-obsoleta, ранее называвшуюся infuscans-obsoleta): обе эти формы имеют нечеткие темные линии под широкими светлыми предплечевыми полосами или вовсе их не имеют.

♀ Фото © Виталий Сирота, Минский р-н
Согласно Скворцову, у самок выделяют 3 основных типа: А, В и С.
А-тип (андроморфные или изоморфные). В ювенильном состоянии имеют лиловую или оранжевую грудь (и основание брюшка) с хорошо выраженной доплечевой полосой. В дальнейшем они становятся ярко-зелеными или голубыми, как самцы, и приобретают характерный для них "фонарь" на конце брюшка.
В-тип (переходные). В ювенильном состоянии имеют лиловую грудь (и начало брюшка), также с хорошо выраженной черной доплечевой полосой. При взрослении они становятся оливково-зелеными или бурыми, без голубого пятна на конце брюшка.
С-тип (гетероморфные или типично гетероморфные). В ювенильном состоянии имеют розовую или оранжевую грудь (и основание брюшка) без черной предплечевой полосы. Во взрослом состоянии они бурые или зеленые, без голубого пятна на конце брюшка.
А-тип (андроморфные или изоморфные). В ювенильном состоянии имеют лиловую или оранжевую грудь (и основание брюшка) с хорошо выраженной доплечевой полосой. В дальнейшем они становятся ярко-зелеными или голубыми, как самцы, и приобретают характерный для них "фонарь" на конце брюшка.
В-тип (переходные). В ювенильном состоянии имеют лиловую грудь (и начало брюшка), также с хорошо выраженной черной доплечевой полосой. При взрослении они становятся оливково-зелеными или бурыми, без голубого пятна на конце брюшка.
С-тип (гетероморфные или типично гетероморфные). В ювенильном состоянии имеют розовую или оранжевую грудь (и основание брюшка) без черной предплечевой полосы. Во взрослом состоянии они бурые или зеленые, без голубого пятна на конце брюшка.

© Виталий Сирота, Минский р-н
От сходного, но гораздо более редкого I. pumilio самцы отличаются голубой меткой на заднем конце брюшка, которая занимает только VIII сегмент, а не часть VIII и весь IX. Общая окраска тела у I. elegans также значительно ярче. У обоих полов птеростигма переднего крыла такого же размера, как и заднего, в отличие от I. pumilio, у которого птеростигма переднего крыла заметно больше. I. elegans также имеет срединную лопасть (отсутствует у I. pumilio) на заднем крае переднеспинки.
Высокая плотность популяции благоприятствует появлению у самок форм, похожих на самцов и ускоряет изменение окраски у других форм. Однако здесь не всё так однозначно, как и объяснение феномена цветового полиморфизма. Далее используются термины: адрохромные - имеющие окраску самцов, гинохромные - все остальные цветовые формы самок.
Ранее рассматривались три теории: 1) репродуктивная изоляция, когда андрохромные самки редко спариваются с самцами, в то время как гинохромные самки делают это; 2) мимикрия под самца, когда самка может оплодотворить запас яиц на всю жизнь за одно спаривание; это позволяет избежать ненужного беспокойства со стороны других самцов (поскольку их можно спутать с самцами), позволяя сосредоточиться на кормлении и откладке яиц; андрохромные самки, будучи крупнее других самок, также могут иметь больше яиц; и 3) при высокой плотности популяции андрохромные самки будут иметь преимущество, поскольку не подвергаются преследованиям со стороны самцов, но будут в невыгодном положении при низкой плотности популяции. Считалось, что теория зависимости от плотности является наиболее вероятным объяснением сохранения полиморфизма окраски самок и что это дает преимущество андрохромным самкам при высокой плотности популяции. Однако это преимущество в уменьшении беспокойства уравновешивается снижением успеха спаривания из-за неспособности самцов распознавать в них самок, в результате чего некоторые андрохромные самки никогда не спарятся.
Высокая плотность популяции благоприятствует появлению у самок форм, похожих на самцов и ускоряет изменение окраски у других форм. Однако здесь не всё так однозначно, как и объяснение феномена цветового полиморфизма. Далее используются термины: адрохромные - имеющие окраску самцов, гинохромные - все остальные цветовые формы самок.
Ранее рассматривались три теории: 1) репродуктивная изоляция, когда андрохромные самки редко спариваются с самцами, в то время как гинохромные самки делают это; 2) мимикрия под самца, когда самка может оплодотворить запас яиц на всю жизнь за одно спаривание; это позволяет избежать ненужного беспокойства со стороны других самцов (поскольку их можно спутать с самцами), позволяя сосредоточиться на кормлении и откладке яиц; андрохромные самки, будучи крупнее других самок, также могут иметь больше яиц; и 3) при высокой плотности популяции андрохромные самки будут иметь преимущество, поскольку не подвергаются преследованиям со стороны самцов, но будут в невыгодном положении при низкой плотности популяции. Считалось, что теория зависимости от плотности является наиболее вероятным объяснением сохранения полиморфизма окраски самок и что это дает преимущество андрохромным самкам при высокой плотности популяции. Однако это преимущество в уменьшении беспокойства уравновешивается снижением успеха спаривания из-за неспособности самцов распознавать в них самок, в результате чего некоторые андрохромные самки никогда не спарятся.

© Оксана С., Минский р-н
Исследование Тиррела (2007) показало, что многие предположения оказались ошибочными. Для этого исследовались 2 различные популяции - с малой плотностью и с большой. В обеих популяциях андрохромные самки составляли более половины зрелой популяции самок, за ними следовали rufescens-obsoleta, а наименее многочисленными были infuscans.
Анализ наблюдений не показал какой-либо статистической разницы в доле каждой цветовой формы между популяциями с высокой и низкой плотностью. Кроме того, что доля размножающихся самок каждой цветовой формы не отличалась от доли этой цветовой формы в популяции в целом. При наблюдении за спариванием самца и андроморфной самки становится особенно ясно, что самка ни в коем случае не является идеальной имитацией самца.
При обеих плотностях популяции поведение каждой цветной формы самок при размножении соответствовало ожидаемой частоте спариваний для популяции в целом. Это означает, что самка не получает никаких преимуществ или недостатков в размножении из-за своей цветовой формы и что самцы легко распознают все цветовые формы самок как своих сородичей.
Действительно, длина тела андрохромных самок значительно больше длины тела гинохромных. Предполагалось, что больший размер тела содержит больше яиц и (или) увеличивает продолжительность жизни. Однако существенной разницы в средней продолжительности жизни между андрохромными и гинохромными самками не обнаружено. Способность выработать повышенное количество яиц не проверялась, однако и предположение было сделано ранее лишь теоретически только по значительно большему размеру тела андрохромных самок.
Анализ наблюдений не показал какой-либо статистической разницы в доле каждой цветовой формы между популяциями с высокой и низкой плотностью. Кроме того, что доля размножающихся самок каждой цветовой формы не отличалась от доли этой цветовой формы в популяции в целом. При наблюдении за спариванием самца и андроморфной самки становится особенно ясно, что самка ни в коем случае не является идеальной имитацией самца.
При обеих плотностях популяции поведение каждой цветной формы самок при размножении соответствовало ожидаемой частоте спариваний для популяции в целом. Это означает, что самка не получает никаких преимуществ или недостатков в размножении из-за своей цветовой формы и что самцы легко распознают все цветовые формы самок как своих сородичей.
Действительно, длина тела андрохромных самок значительно больше длины тела гинохромных. Предполагалось, что больший размер тела содержит больше яиц и (или) увеличивает продолжительность жизни. Однако существенной разницы в средней продолжительности жизни между андрохромными и гинохромными самками не обнаружено. Способность выработать повышенное количество яиц не проверялась, однако и предположение было сделано ранее лишь теоретически только по значительно большему размеру тела андрохромных самок.

© Виталий Сирота, г. Бобруйск, Могилевская обл.
Имеет широкий диапазон местообитаний, населяет почти все типы стоячих и проточных водоемов, но не любит слишком кислых и слишком быстрых вод: озера, пруды, канавы, глубокие лужи, болотные ямы и мочажины, медленно текущие речки, ручьи, старицы с богатой водной растительностью. Размер водоема не имеет решающего значения, вид встречается как в крупных озерах, так и в небольших прудах, которые часто имеют антропогенное происхождение, например, бетонные бассейны. Водоемы начальных стадий (без растительности) и водоемы с эфемерным течением исключают появление вида. Переносит солоноватую и загрязненную воду. Из-за приспособляемости к среде обитания может встречаться с большинством других видов стрекоз, включая I. pumilio.
Вид, вероятно, имеет хороший потенциал для экспансии, так как вновь созданные водоемы заселяются даже на большем расстоянии от исходного местообитания спустя относительно короткое время с момента их создания, как только появляется разреженная береговая или водная растительность. Это объясняет большое количество регистраций в бывших карьерах.
Большинство взрослых особей держатся близко к воде, нередко собираясь в больших количествах среди маргинальной растительности. Часто присутствует у воды, когда плохая погода заставляет другие виды держаться вдали. В отличие от других видов, пик численности не всегда приходится на полдень. Взрослые насекомые часто устраиваются на ночлег у воды. Они выбирают для ночлега узкие стебли, что связано с размерами головы: поэтому их часто можно найти там, где растет тростник.
Максимальная продолжительность жизни, наблюдаемая в полевых условиях, составляет 45 дней. Существует вероятность, что некоторые особи могут прожить около 50 дней.
Несмотря на слабый полет, охотно расселяется на новые места.
Вид, вероятно, имеет хороший потенциал для экспансии, так как вновь созданные водоемы заселяются даже на большем расстоянии от исходного местообитания спустя относительно короткое время с момента их создания, как только появляется разреженная береговая или водная растительность. Это объясняет большое количество регистраций в бывших карьерах.
Большинство взрослых особей держатся близко к воде, нередко собираясь в больших количествах среди маргинальной растительности. Часто присутствует у воды, когда плохая погода заставляет другие виды держаться вдали. В отличие от других видов, пик численности не всегда приходится на полдень. Взрослые насекомые часто устраиваются на ночлег у воды. Они выбирают для ночлега узкие стебли, что связано с размерами головы: поэтому их часто можно найти там, где растет тростник.
Максимальная продолжительность жизни, наблюдаемая в полевых условиях, составляет 45 дней. Существует вероятность, что некоторые особи могут прожить около 50 дней.
Несмотря на слабый полет, охотно расселяется на новые места.

Ночующие особи разных полов Фото © Максим Мясников, Дзержинский р-н, Минская обл.
Самцы территориальны. Например, в Британии самая высокая постоянная плотность составляет около 30 самцов на 100 м уреза воды, хотя они могут встречаться в гораздо большем количестве в близлежащей маргинальной растительности. При взаимодействии с другими стрекозами быстро дергается вверх и вниз в полете.
При первом спаривании особи могут еще не достичь зрелой окраски. Зарегистрировано, что созревание rufescens в rucfescens-obsoleta (ранее называвшееся infuscans-obsoleta) занимает в среднем 8,1 дня, тогда как созревание violacea в infuscans занимает всего 6,9 дня. Если предположить, что половая зрелость достигается в одно и то же время в обоих случаях, то либо rufescens становится зрелой до того, как становится rufescens-obsoleta, либо violacea остается незрелой до тех пор, пока не становится infuscans; это также может быть причиной того, что единственными «неполовозрелыми» самками, часто наблюдаемыми спаривающимися, бывают rufescens. Однако следует отметить, что I. elegans может спариваться всего через 3-4 дня, так что, возможно, обе неполовозрелые формы становятся половозрелыми до того, как они меняют окраску.
Совокупление длится дольше, чем у любой другой стрекозы: от 2 до 6 часов, и обычно достигает пика во второй половине дня, после чего самки могут откладывать яйца до вечера.
Яйца откладываются в ткани водных растений и растительный мусор на поверхности воды. Данные о присутствии самца при откладке яиц различаются. Согласно старым советским источникам, самец обычно сопровождает самку при кладке яиц, которая иногда во время этого процесса опускает дистальную половину брюшка в воду, но никогда не спускается глубже. Согласно британским и немецким авторам, яйцекладка без присутствия самцов.
При первом спаривании особи могут еще не достичь зрелой окраски. Зарегистрировано, что созревание rufescens в rucfescens-obsoleta (ранее называвшееся infuscans-obsoleta) занимает в среднем 8,1 дня, тогда как созревание violacea в infuscans занимает всего 6,9 дня. Если предположить, что половая зрелость достигается в одно и то же время в обоих случаях, то либо rufescens становится зрелой до того, как становится rufescens-obsoleta, либо violacea остается незрелой до тех пор, пока не становится infuscans; это также может быть причиной того, что единственными «неполовозрелыми» самками, часто наблюдаемыми спаривающимися, бывают rufescens. Однако следует отметить, что I. elegans может спариваться всего через 3-4 дня, так что, возможно, обе неполовозрелые формы становятся половозрелыми до того, как они меняют окраску.
Совокупление длится дольше, чем у любой другой стрекозы: от 2 до 6 часов, и обычно достигает пика во второй половине дня, после чего самки могут откладывать яйца до вечера.
Яйца откладываются в ткани водных растений и растительный мусор на поверхности воды. Данные о присутствии самца при откладке яиц различаются. Согласно старым советским источникам, самец обычно сопровождает самку при кладке яиц, которая иногда во время этого процесса опускает дистальную половину брюшка в воду, но никогда не спускается глубже. Согласно британским и немецким авторам, яйцекладка без присутствия самцов.

© Виталий Сирота, Минский р-н
Нимфа. Окраска тела сильно варьирует, встречаются личинки желтоватые, коричневатые, зеленоватые, зеленовато-серые; тело слегка удлиненное, длиной 23-26 мм, покрыто шипиками. Голова широкая, 3,5-4 мм в поперечнике; темя слегка выпуклое, с хорошо заметными глазками; глаза большие, выпуклые, не выдающиеся над затылком; затылок узкий, с сильно скошенными внешними сторонами, с округлыми задними углами, покрытыми шипиками, с сильно вогнутым задним краем. Маска в состоянии покоя простирается до основания средней пары ног; подбородок с очень узким основанием и широким дистальным краем, длиной 2,2 мм. Переднегрудь короткая, угловатая по бокам, с темными краями и темными небольшими продольными и поперечными линиями. Крыловые чехлы довольно короткие, простираются до основания IV сегмента На бедрах по 1 темному кольцу близ дистального края; бедра и голени покрыты шипиками. Брюшко со светлой дорсальной срединной полоской и светлыми краями; задний край Х сегмента снабжен маленькими шипиками; межсегментальные пространства светлые с темными пятнами; вентральная сторона светлее, чем дорсальная; латеральные шипы на V-IX сегментах брюшка маленькие, острые. Хвостовые жаберные пластинки очень длинные, ланцетовидные, довольно узкие, приблизительно в 5 раз длиннее своей ширины, с длинным острием на вершине; деление на 2 части: базальную (более узкую) и дистальную (более широкую), заметно по более или менее слабому следу поперечного узелкового шва и по окончанию ряда предузелковых краевых шипов; последние на вентральном крае пластинок толще, сидят плотнее и простираются приблизительно в 2,5 раза дальше, чем на дорсальном, где они расположены лишь при основании; боковые края дистальной части снабжены тонкими короткими волосками; средний трахеальный ствол большею частью приближен к вентральной стороне, многочисленные разветвления трахей темные. Боковые створки яйцеклада самки простираются до половины Х сегмента.

Фото © Денис Китель.
Личинки неприхотливы в выборе места обитания и живут в самых разнообразных водоемах, как со стоячей водой, даже загрязненной и солоноватой, так и в водах проточных, с медленным или довольно сильным течением, в местах, богатых водной растительностью и не особенно глубоких (0,1-0,4 м). К загрязнению воды терпимы. Развитие личинок в течение 1 или 2 лет, в зависимости от широты. Ранее самое короткое время от откладки яиц до появления имаго составляло 63 дня, зарегистрированное на греческих рисовых полях. Однако данные из Верхнего Рейна в районе Карлсруэ, где затопление кукурузного поля, начавшееся очень холодной весной и продолжавшееся до начала лета, показали, что потребовалось 55 дней для развития яиц и личинок от первого дня затопления (13.05.2016 г.) до первого наблюдаемого вылета (07.07.2016 г.). Из-за холодной и дождливой погоды в начале затопления первая колонизация видом не могла произойти в первые сутки после затопления. Поэтому предполагается, что вероятен еще более короткий период развития.
Имаго иногда (чаще, чем другие стрекозы) отрождаются с экзувиями, направленными головой вниз. В отличие от других стрекоз Zygoptera, неполовозрелые особи часто встречаются у кромки воды. Хотя по другим данным, лишь при отрождении все стрекозы находятся у воды. Сразу после этого их количество падает, но большая часть популяции возвращается на 10-й день. Время созревания составляет у самцов 10 дней, у самок - 12 дней.
После основного периода отрождения имаго в начале лета встречи экзувий и только что вылупившихся особей в августе уже редки. Возобновление отрождения в конце лета, иногда даже в сентябре, указывает на цикл развития личинок в течение года и формирование второго поколения. В то же время, заметного увеличения позднеосенних находок в последние годы не наблюдается, что указывает на то, что возможность второго поколения за год не связана с глобальным потеплением, а зависит от конкретных условий определенной локации и определенного года.
Имаго иногда (чаще, чем другие стрекозы) отрождаются с экзувиями, направленными головой вниз. В отличие от других стрекоз Zygoptera, неполовозрелые особи часто встречаются у кромки воды. Хотя по другим данным, лишь при отрождении все стрекозы находятся у воды. Сразу после этого их количество падает, но большая часть популяции возвращается на 10-й день. Время созревания составляет у самцов 10 дней, у самок - 12 дней.
После основного периода отрождения имаго в начале лета встречи экзувий и только что вылупившихся особей в августе уже редки. Возобновление отрождения в конце лета, иногда даже в сентябре, указывает на цикл развития личинок в течение года и формирование второго поколения. В то же время, заметного увеличения позднеосенних находок в последние годы не наблюдается, что указывает на то, что возможность второго поколения за год не связана с глобальным потеплением, а зависит от конкретных условий определенной локации и определенного года.

♀ Фото © Семён Левый, Кобринский р-н, Брестская обл.

♂ Фото © Семён Левый, Кобринский р-н, Брестская обл.

© Виталий Сирота

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Common Bluetail. Ischnura elegans" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 106-107.
9. Smallshire D., Swash A. "Blue-tailed Damselfly. Ischnura elegans" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 92-94.
10. Hinnekint B. O. N. "Population dynamics of Ischnura E. elegans (Vander Linden) (Insecta: Odonata) with special reference to morphological colour changes, female polymorphism, multiannual cycles and their influence on behaviour" / Hydrobiologia 146 : 3-31 (1987)
11. Martens A., Dunst J., Fröhlich A., Grabow K. "Just 55 days: Rapid development of Ischnura elegans and I. pumilio in a flooded maize field in Central Europe (Odonata: Coenagrionidae)" / Notulae odonatologicae 8(10) 2017: 369-392.
12. Tyrrel M. "Maintenance of the female androchrome colour polymorph in the Blue-tailed Damselfly Ischnura elegans (Vander Linden)" / Journal of the British Dragonfly Society, Volume 23 No. 2, 2007 33-39.
13. Lehmann A., Lehmann G., Brauner O. "Große Pechlibelle – Ischnura elegans (Vander Linden 1820)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 56-57.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Common Bluetail. Ischnura elegans" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 106-107.
9. Smallshire D., Swash A. "Blue-tailed Damselfly. Ischnura elegans" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 92-94.
10. Hinnekint B. O. N. "Population dynamics of Ischnura E. elegans (Vander Linden) (Insecta: Odonata) with special reference to morphological colour changes, female polymorphism, multiannual cycles and their influence on behaviour" / Hydrobiologia 146 : 3-31 (1987)
11. Martens A., Dunst J., Fröhlich A., Grabow K. "Just 55 days: Rapid development of Ischnura elegans and I. pumilio in a flooded maize field in Central Europe (Odonata: Coenagrionidae)" / Notulae odonatologicae 8(10) 2017: 369-392.
12. Tyrrel M. "Maintenance of the female androchrome colour polymorph in the Blue-tailed Damselfly Ischnura elegans (Vander Linden)" / Journal of the British Dragonfly Society, Volume 23 No. 2, 2007 33-39.
13. Lehmann A., Lehmann G., Brauner O. "Große Pechlibelle – Ischnura elegans (Vander Linden 1820)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 56-57.
