СТРЕЛКА МАЛЕНЬКАЯ или ИШНУРА МАЛЕНЬКАЯ
Ischnura pumilio Charpentier, 1825

♂ Фото © Виктор Рындевич, г. Минск
Ischnura pumilio Charpentier, 1825 (стрелка (ишнура) маленькая)
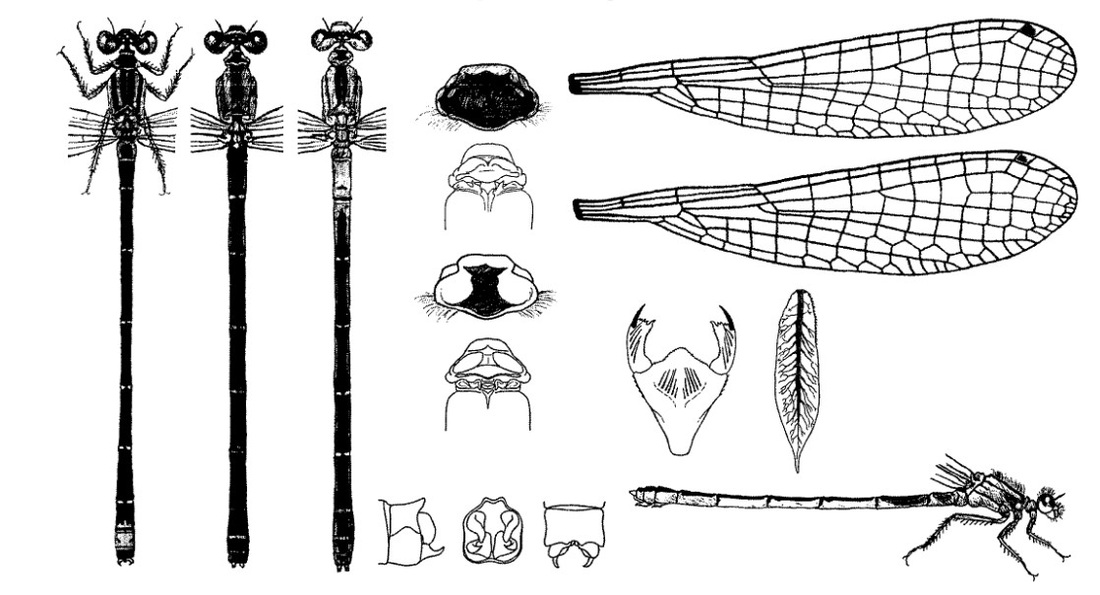
Голова широкая голова, сверху вдвое шире своей длины. Лоб округлен. На затылке 2 светлых пятна. Затылочные пятна округленные. Задний край переднеспинки несколько вытянут назад, простой, не дифференцированный. Птеростигмы двухцветные, узкие (равны 1 ячейке), менее чем в 2 раза длиннее ширины, на передних крыльях заметно крупнее, чем на задних, особенно у самцов. Ноги черные или темно-серые.
♂: Верхняя часть глаз черная, нижняя голубая или голубовато-зеленая; такого же цвета как низ глаз бока груди и узкие предплечевые полосы. Сверху грудь черная. Брюшко сверху темное с характерным голубым "фонарем" (VIII сегмент частично голубой (задняя треть), IX сегмент целиком голубой), с боков и снизу голубоватое или несколько первых сегментов зеленоватые. На сегменте IX обычно имеет пару крошечных черных пятен на дорсальной стороне, хотя иногда они увеличены. Нижняя сторона брюшка желтоватая или зеленоватая. Птеростигма на передних крыльях ясно двухцветная, с черной проксимальной частью и беловатой дистальной, на задних другой формы и значительно меньше.
♀: Глаза зеленоватые. Бока груди также зеленоватые. Широкие буровато-зеленые предплечевые полосы с очень узкой черной полосой внизу. Изредка встречается голубая форма (андроморфная). Брюшко сверху черное, без светлого сегмента VIII (у обеих форм). На VIII стерните брюшка перед яйцекладом острый шип. Анальные придатки белые, иногда лишь у основания черноватые. Птеростигмы более коричневые и менее отчетливо двухцветные, чем у самца.
У неполовозрелых самцов окрашенные части бледнее и зеленее. Неполовозрелые самки интенсивно оранжевые, оранжевый цвет доходит до жилок крыльев и вершин сегментов I-III. Глаза, ноги, бока груди и брюшко ярко-оранжевого цвета. Они известны как var. aurantiaca.
Длина 26-31 мм (26–30 мм у самцов и 28–31 мм у самок), брюшко 22-25 мм, заднее крыло 14-18 мм.
Лет: конец мая - середина октября. Возможно, имеет 2 пика, в июне и августе.
♂: Верхняя часть глаз черная, нижняя голубая или голубовато-зеленая; такого же цвета как низ глаз бока груди и узкие предплечевые полосы. Сверху грудь черная. Брюшко сверху темное с характерным голубым "фонарем" (VIII сегмент частично голубой (задняя треть), IX сегмент целиком голубой), с боков и снизу голубоватое или несколько первых сегментов зеленоватые. На сегменте IX обычно имеет пару крошечных черных пятен на дорсальной стороне, хотя иногда они увеличены. Нижняя сторона брюшка желтоватая или зеленоватая. Птеростигма на передних крыльях ясно двухцветная, с черной проксимальной частью и беловатой дистальной, на задних другой формы и значительно меньше.
♀: Глаза зеленоватые. Бока груди также зеленоватые. Широкие буровато-зеленые предплечевые полосы с очень узкой черной полосой внизу. Изредка встречается голубая форма (андроморфная). Брюшко сверху черное, без светлого сегмента VIII (у обеих форм). На VIII стерните брюшка перед яйцекладом острый шип. Анальные придатки белые, иногда лишь у основания черноватые. Птеростигмы более коричневые и менее отчетливо двухцветные, чем у самца.
У неполовозрелых самцов окрашенные части бледнее и зеленее. Неполовозрелые самки интенсивно оранжевые, оранжевый цвет доходит до жилок крыльев и вершин сегментов I-III. Глаза, ноги, бока груди и брюшко ярко-оранжевого цвета. Они известны как var. aurantiaca.
Длина 26-31 мм (26–30 мм у самцов и 28–31 мм у самок), брюшко 22-25 мм, заднее крыло 14-18 мм.
Лет: конец мая - середина октября. Возможно, имеет 2 пика, в июне и августе.

♀ Фото © Виктор Рындевич, Минский р-н.
Самцы меньше самок. Неполовозрелая форма имеет окраску, отличную от зрелой формы, с VIII и IX сегментами брюшка, окрашенными в коричневый или желто-серый цвет. Эта стадия длится менее 48 часов, прежде чем появляются признаки синевы. Однако имеется наблюдение, как три выращенных в неволе самца становились оранжевыми, как aurantiaca, переходя в промежуточную желто-серую стадию в течение 24 часов и становясь типичными взрослыми особями с темным брюшком и голубым хвостом в течение следующих 24 часов. Иногда промежуточные формы незрелой и зрелой окраски наблюдаются как у самцов, так и у самок, но их редкость у самцов указывает на то, что изменение окраски происходит быстро.
Только полностью развитая форма (типичная и андроморфная) у самок имеет крупные зрелые яйца. В некоторых исследованиях самки aurantiaca не наблюдались копулирующими и казались непривлекательными для самцов. В других исследованиях наблюдалось влечение самцов к этой форме, что приводило к энергичным попыткам отвергнуть самцов самками aurantiaca на ранней стадии развития и некоторым совокуплениям самок этой формы на поздней стадии. Последнее предполагает, что сперматозоиды могут сохраняться до тех пор, пока яйцеклетки не созреют, так как самки aurantiaca никогда не наблюдались откладывающими яйца.
Только полностью развитая форма (типичная и андроморфная) у самок имеет крупные зрелые яйца. В некоторых исследованиях самки aurantiaca не наблюдались копулирующими и казались непривлекательными для самцов. В других исследованиях наблюдалось влечение самцов к этой форме, что приводило к энергичным попыткам отвергнуть самцов самками aurantiaca на ранней стадии развития и некоторым совокуплениям самок этой формы на поздней стадии. Последнее предполагает, что сперматозоиды могут сохраняться до тех пор, пока яйцеклетки не созреют, так как самки aurantiaca никогда не наблюдались откладывающими яйца.

♀ Фото © Виктор Рындевич, г. Минск
Андроморфная (или гетерохромная) форма самок, как правило, встречается редко, например, в Великобритании о ней вообще не сообщалось. Исследование в Германии показало, что самки-андроморфы всегда менее многочисленны, чем гиноморфы, и были обнаружены только в 6 из 10 посещенных мест.
Андроморфные самки претерпевают на одно изменение цвета больше; сначала они меняют с оранжевую окраску aurantiaca на типичную зеленую, а затем с зеленой на лазурно-голубую, типичную для самцов. Эти два изменения цвета часто сливаются, и можно найти особей, демонстрирующих все три типа окраски. Зеленых самок можно идентифицировать как будущих андроморфных по степени меланизма на различных сегментах брюшка, но в настоящее время невозможно выделить окончательный фенотип самок aurantiaca.
Вместе с тем, хотя подавляющее количество специалистов еще в конце ХХ в. пришло к выводу, что aurantiaca является ювенильной формой, еще остаются авторы, считающие ее отдельной морфой взрослых самок. Например, впервые для Норвегии обнаруженные в самом начале июня самки указываются как взрослые гетерохромные и уникальные для Норвегии тем, что это единственные стрекозы семейства с оранжевой окраской. Впрочем, дальнейшие наблюдения не велись, что и привело к таким довольно неоднозначным выводам (наблюдения с 30.05 по 02.06.2012, Бергсдален, Норвегия).
Андроморфные самки претерпевают на одно изменение цвета больше; сначала они меняют с оранжевую окраску aurantiaca на типичную зеленую, а затем с зеленой на лазурно-голубую, типичную для самцов. Эти два изменения цвета часто сливаются, и можно найти особей, демонстрирующих все три типа окраски. Зеленых самок можно идентифицировать как будущих андроморфных по степени меланизма на различных сегментах брюшка, но в настоящее время невозможно выделить окончательный фенотип самок aurantiaca.
Вместе с тем, хотя подавляющее количество специалистов еще в конце ХХ в. пришло к выводу, что aurantiaca является ювенильной формой, еще остаются авторы, считающие ее отдельной морфой взрослых самок. Например, впервые для Норвегии обнаруженные в самом начале июня самки указываются как взрослые гетерохромные и уникальные для Норвегии тем, что это единственные стрекозы семейства с оранжевой окраской. Впрочем, дальнейшие наблюдения не велись, что и привело к таким довольно неоднозначным выводам (наблюдения с 30.05 по 02.06.2012, Бергсдален, Норвегия).

♂ Фото © Денис Китель
Созревание у самки занимает несколько больше времени, чем у самца: стадия aurantiaca длится 5-9 дней, промежуточная стадия - 2 дня, а зрелая самка живет еще до 26 дней. В неволе для созревания требуется 6-12 дней, а для изменения цвета обычно 2 дня. Время созревания обратно пропорционально количеству пищи и температуре. Однако дата отрождения имаго очень сильно коррелирует со временем созревания: более позднее отрождение приводит к более короткому времени до изменения цвета.
В лабораторных условиях общая продолжительность жизни имаго у самок значительно больше, чем у самцов (самки 16,3±1,94 и максимум 37 дней; самцы 10,7±1,02 и максимум 26 дней). Одна самка, исключенная из анализа, прожила 57 дней. Напротив, полевые исследования показывают, что продолжительность жизни самок меньше, чем у самцов. Однако высказано предположение, что реальной разницы не существует и что предполагаемая разница связана с повышенной вероятностью повторной поимки самцов по сравнению с самками.
В лабораторных условиях общая продолжительность жизни имаго у самок значительно больше, чем у самцов (самки 16,3±1,94 и максимум 37 дней; самцы 10,7±1,02 и максимум 26 дней). Одна самка, исключенная из анализа, прожила 57 дней. Напротив, полевые исследования показывают, что продолжительность жизни самок меньше, чем у самцов. Однако высказано предположение, что реальной разницы не существует и что предполагаемая разница связана с повышенной вероятностью повторной поимки самцов по сравнению с самками.

♂ Фото © Виктор Рындевич, Минский р-н.
Места обитания - почти все типы стоячих или медленно текущих водоемов, чаще небольших, как временных, так и непересыхающих. Предпочитает скудно заросшие, теплые, мелкие, часто эфемерные воды. В Скандинавии отмечается, что использование бывших карьеров важно в качестве мест размножения. Вероятно, это связано с благоприятным температурным режимом в таких местах в сочетании с мелководьем и малой численностью растительности. Как вид, основной ареал которого находится южнее, вероятно, может переносить частичное высыхание, но не промерзание. Это выражается, в частности, в том, что места с выходом подземных вод (т. е. не промерзающие) для вида явно предпочтительнее.
Считается, что у вида очень специфические требования к среде обитания, что является причиной его очевидной редкости. Вид ограничен мелководьем, водой с медленным течением и требует определенной степени открытости в виде голой, илистой земли и редкой растительности. В этом специфическом микроклимате вид обитает в самых разных природных и искусственных местах. Был зарегистрирован в естественных местообитаниях, таких как заболоченные старицы, берега луж и ручьев, а также болота. Быстро заселяет вновь образовавшуюся среду обитания, такую как рвы, карьеры и заболоченные земли, образовавшиеся в результате добычи глины, гравия, песка, известняка и мела. Был зарегистрирован в прудах для сбора ливневых вод на автомагистралях во Франции, которые содержали высокие концентрации меди и цинка в верхнем слое отложений и гербицидов в толще воды. Отмечена откладка яиц в луже на обочине дороги.
Отсутствует в наиболее кислых местообитаниях, например, в тех, где преобладает сфагнум.
Считается, что у вида очень специфические требования к среде обитания, что является причиной его очевидной редкости. Вид ограничен мелководьем, водой с медленным течением и требует определенной степени открытости в виде голой, илистой земли и редкой растительности. В этом специфическом микроклимате вид обитает в самых разных природных и искусственных местах. Был зарегистрирован в естественных местообитаниях, таких как заболоченные старицы, берега луж и ручьев, а также болота. Быстро заселяет вновь образовавшуюся среду обитания, такую как рвы, карьеры и заболоченные земли, образовавшиеся в результате добычи глины, гравия, песка, известняка и мела. Был зарегистрирован в прудах для сбора ливневых вод на автомагистралях во Франции, которые содержали высокие концентрации меди и цинка в верхнем слое отложений и гербицидов в толще воды. Отмечена откладка яиц в луже на обочине дороги.
Отсутствует в наиболее кислых местообитаниях, например, в тех, где преобладает сфагнум.

♂ Фото © Виталий Сирота, Минск
Ручьи, занятые видом, обычно представляют собой богатые щелочами потоки, в которых иногда преобладают камыши Juncus и часто обитают Orthetrum coerulescens, Pyrrhosoma nymphula, Sympetrum striolatum и Cordulegaster boltonii. В прудах P. nymphula и S. striolatum являются обычными спутниками этого вида, и к ним часто присоединяются I. elegans, Enallagma cyathigerum, Coenagrion puella, Anax imperator и Libellula depressa. Хотя чрезмерный рост растений может исключить размножение вида, для откладывания яиц требуется некоторая надводная растительность, обычно с мягкими стеблями. Типичные встречающиеся виды растений включают виды Juncus, виды Typha, Agrostis stolonifera, Glyceria fluitans и Ranunculus flammula. Считается, что тенеральные особи и взрослые самки укрываются в густых зарослях болотного мирта.

♀ Фото © Денис Китель, Малоритский р-н, Брестская обл.
Хрупкая небольшая стрекоза, незаметная и легко упускаемая из виду, полет слабый, трепещущий, обычно низко передвигается среди травы и других прибрежных растений, где иногда и отдыхают. Несмотря на слабый внешний вид, рассеивается на большие расстояния, используя благоприятные воздушные потоки, когда плотность растений достигает критической стадии. Например, в прибрежных районах Великобритании иногда регистрируются иммигранты из континентальной Европы. Как вид-первопроходец на новых участках, очень подвижен, а его популяции часто недолговечны. Популяции могут быть довольно плотными.
Одной из причин отсутствия знаний о биологии вида является временный характер его колоний. Вид быстро заселяет вновь образованные биотопы, но колонии, как правило, вымирают через несколько лет. Часто встречается в местах, удаленных от ближайшей известной колонии. Это говорит о том, что у него есть возможность рассеяться на значительное расстояние, что не соответствует наблюдаемому слабому беспорядочному полету. Однако был обнаружен потенциальный метод рассредоточения на большие расстояния. В нескольких случаях отдельные зрелые взрослые самцы демонстрировали сильный взлет вверх на высоту, на которой они больше не были видны в бинокль.
Наблюдения происходили в ясные, тихие и очень жаркие дни, что могло способствовать развитию сильных восходящих потоков воздуха. Возможно, стрекозы использовали для облегчения полета вверх, чтобы затем, с помощью направленных потоков воздуха перелетать над значительными препятствиями, такими как деревья и здания. Более сильных порывов на большей высоте может быть достаточно для рассредоточения на большие расстояния. Такое сильное целенаправленное расселение необычно для половозрелых стрекоз и может давать им возможность размножаться до неопределенного исхода расселения.
Одной из причин отсутствия знаний о биологии вида является временный характер его колоний. Вид быстро заселяет вновь образованные биотопы, но колонии, как правило, вымирают через несколько лет. Часто встречается в местах, удаленных от ближайшей известной колонии. Это говорит о том, что у него есть возможность рассеяться на значительное расстояние, что не соответствует наблюдаемому слабому беспорядочному полету. Однако был обнаружен потенциальный метод рассредоточения на большие расстояния. В нескольких случаях отдельные зрелые взрослые самцы демонстрировали сильный взлет вверх на высоту, на которой они больше не были видны в бинокль.
Наблюдения происходили в ясные, тихие и очень жаркие дни, что могло способствовать развитию сильных восходящих потоков воздуха. Возможно, стрекозы использовали для облегчения полета вверх, чтобы затем, с помощью направленных потоков воздуха перелетать над значительными препятствиями, такими как деревья и здания. Более сильных порывов на большей высоте может быть достаточно для рассредоточения на большие расстояния. Такое сильное целенаправленное расселение необычно для половозрелых стрекоз и может давать им возможность размножаться до неопределенного исхода расселения.

♀ Фото © Виталий Сирота, Минский р-н.
Хотя считается «бродячим оппортунистом» из-за склонности появляться в местах, удаленных от известных местонахождений, однако это касается преимущественно временных местообитаний. Предполагается, что перемещение на большие расстояния из основных местообитаний происходит редко. Среднее перемещение за всю жизнь 2304 повторно отловленных помеченных особей в Великобритании, составило 56 м, а максимальное перемещение составило 1165 м. Это означает, по-видимому, что вид является одним из малоподвижных. Быстрое и дальнее распространение этого вида, вероятно, является случайным, если оно происходит из основных местообитаний, и, вероятно, является результатом сочетания благоприятного климата в последние годы в сочетании с увеличением популяции, занимающей субоптимальные или временные локации. Спаривающиеся самки, унесенные сильным южным ветром, могут случайно оказаться рядом с потенциальным местом размножения и затем легко начать новую популяцию.
Самцы не территориальны, но конкурируют за доступ к самкам. Исследование успеха спаривания в течение жизни показало, что большая часть самцов никогда не спаривается (43,6%) по сравнению с небольшим числом самок (13%). Разброс успешности был выше у самцов; один самец достиг максимума 7 спариваний, тогда как максимум для самки был 4 спаривания. Особи с большей продолжительностью жизни достигают наибольшего количества спариваний, но размер тела не оказал существенного влияния на успешность ни у одного из полов. Продолжительность копуляции в среднем составляла 1 час 19 минут, но одна пара наблюдалась в копуле около 5 час. Продолжительность копуляции отрицательно коррелирует со временем суток.
Самка откладывает яйца в одиночку, они выше или чуть ниже уровня воды. Откладывание каждого яйца занимает примерно 20 сек. Для кладки используются различные виды растений с мягкими стеблями, в том числе Juncus inflexus, J. articulatus и Eleocharis palustris. В Британии наблюдалась откладка яиц в ситник и харовые водоросли. Там, где имелась одинаковая плотность этих растений, было замечено, что более 75% самок откладывают яйца в J. articulatus, что указывает на предпочтение определенных видов, когда их несколько. Самки откладывают яйца вниз по стеблю, пока грудь не соприкоснется с водой, но попыток полного погружения не зарегистрировано, возможно, из-за проблем с преодолением поверхностного натяжения. Способность вида сохраняться в водоемах, подверженных высыханию летом, может быть связана с откладкой яиц в определенные части стебля, где яйца находятся в растительной ткани и защищены от высыхания.
Яйца вылупляются примерно через 17 дней, хотя при засухе вылупление откладывается до благоприятного момента.
Яйца вылупляются примерно через 17 дней, хотя при засухе вылупление откладывается до благоприятного момента.
Нимфа. Окраска тела желтовато-зеленая; тело гладкое; длина, включая хвостовые жаберные пластинки, 21-22 мм. Голова плоская, в ширину больше, чем в длину; глазки отчетливо выступают в виде бледных светлых пятнышек; 1-й членик антенн самый длинный, его основание и верхушка окольцованы черным, у других члеников только верхушки темные; задние углы затылка тупо округлые; внешние и задние стороны его несут 5-7 маленьких шипиков, задний край сильно вогнут. Подбородок маски напоминает таковой же у I. elegans, но значительно меньших размеров, приблизительно 1,25 мм. Переднегрудь короткая, угловатая по бокам, со светлой линией посредине, с темными краями и темными небольшими продольными и поперечными линиями. Крыловые чехлы достигают IV сегмента брюшка, имеют несколько цилиндрическую или колбасовидную форму. Ноги голые, лишенные крупных шипов, на бедрах несколько небольших щетинок; бедра задней пары ног простираются до уровня верхушек крыловых чехлов; на всех бедрах по срединному и дистальному темному кольцу; на голенях у проксимального конца по одному темному кольцу; дистальный конец голеней передней пары ног на внутренней стороне обрамлен пучком небольших щетинок, образующих как бы щеточку, при помощи которых личинки очищают ротовое отверстие от посторонних предметов. Брюшко с узкой бледной продольной дорсальной центральной полосой, с каждой стороны которой проходят двойные темные полосы, расширяющиеся и сливающиеся у основания каждого сегмента, за исключением Х сегмента; межсегментные пространства светлые с темными полосками и пятнами; латеральные шипы на IV-IX сегментах маленькие, острые. Хвостовые жаберные пластинки длинные (5,0-5,5 мм), узкие, ланцетовидные, с длинным острием на вершине; на поверхности жабер нет видимых следов, поперечного шва, слабые следы можно заметить на месте окончания предузелковых шипов.
Личинки держатся среди водной растительности, очень пугливы и при малейшей опасности быстро удаляются на дно. Личинки этого вида обитают в тех же водоемах, где и личинки I. elegans; экология и биология обоих видов похожи. Личинки обычно развиваются в течение одного года, но два поколения в год являются нормальным явлением в Южной Европе и иногда могут встречаться севернее. Например, в Германии доказаны моно- и бивольтинные циклы развития, т. е. с зимующим поколением и «быстрым» летним поколением. Имеются свидетельства того, что развитие личинок происходит и зимой, что имеет решающее значение для его успешного завершения.
Однако не все выращиваемые в неволе личинки завершают свой жизненный цикл в течение года. В лабораторных условиях 90% предличинок вылупились в одно и то же утро после 17 дней при 25°С, а остальные - на следующий день. В условиях засухи предличинки вылупились через 2 дня после повторного погружения растения в воду.
После вылупления предличинки линяют и появляется личинка 2-го возраста. В течение нескольких минут они выказывают угрозу друг другу, что, по-видимому, побуждает их рассредоточиться от переполненного стебля. Это необычно для стрекоз, у других видов проявления угрозы наблюдаются только с 4-го возраста. Раннее расселение может быть выгодным в условиях мелководья, поскольку в случае засухи некоторые личинки выживут на участках, которые остаются влажными.
Однако не все выращиваемые в неволе личинки завершают свой жизненный цикл в течение года. В лабораторных условиях 90% предличинок вылупились в одно и то же утро после 17 дней при 25°С, а остальные - на следующий день. В условиях засухи предличинки вылупились через 2 дня после повторного погружения растения в воду.
После вылупления предличинки линяют и появляется личинка 2-го возраста. В течение нескольких минут они выказывают угрозу друг другу, что, по-видимому, побуждает их рассредоточиться от переполненного стебля. Это необычно для стрекоз, у других видов проявления угрозы наблюдаются только с 4-го возраста. Раннее расселение может быть выгодным в условиях мелководья, поскольку в случае засухи некоторые личинки выживут на участках, которые остаются влажными.
Личинки живут в или на наносных и илистых субстратах, связанны с ранними сукцессионными растениями.
Важными особенностями микросреды обитания являются мелководье (как стоячее, так и с медленным течением), с теплой водой круглый год. Важны илистый субстрат и минимальная растительность, а также низкий уровень тени. Могут переносить воду разного качества, кислую и щелочную, и даже переносить слегка солоноватую.
На отрождение имаго влияет температура, обычно оно продолжается с конца мая до конца июня. Отрождение имаго высоко синхронизировано как в дикой природе, так и в лаборатории. Половой зрелости вид достигает не у воды, а появляется здесь в большом количестве лишь во взрослом состоянии.
Важными особенностями микросреды обитания являются мелководье (как стоячее, так и с медленным течением), с теплой водой круглый год. Важны илистый субстрат и минимальная растительность, а также низкий уровень тени. Могут переносить воду разного качества, кислую и щелочную, и даже переносить слегка солоноватую.
На отрождение имаго влияет температура, обычно оно продолжается с конца мая до конца июня. Отрождение имаго высоко синхронизировано как в дикой природе, так и в лаборатории. Половой зрелости вид достигает не у воды, а появляется здесь в большом количестве лишь во взрослом состоянии.

♀ Фото © Виталий Сирота, Минский р-н

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
6. Smallshire D., Swash A. "Scarce Blue-tailed Damselfly. Ischnura pumilio" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 94-96.
7. Smallshire D., Swash A. "Small Bluetail. Ischnura pumilio" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 102-103.
8. Moreno-Benítez J. M., Ripoll J. "Ischnura pumilio" / in: Dragonflies along the Great Málaga Path and in the Province. Identification Guide, 2018. pp. 72-73.
9. Olberg S., Lønnve O. J. "Ischnura pumilio (Charpentier, 1825) (Odonoata, Coenagrionidae) in Norway" / Norwegian Journal of Entomology 59, 229–233. 2012.
10. Cham S., Nelson B., Parr A., Prentice S., Smallshire D., Taylor, P. "Atlas of the Dragonflies in Britain and Ireland". Centre for Ecology & Hydrology, 2014. - 288 pp.
11. Allen K. A., Le Duc M. G., Thompson D. J. "Habitat and conservation of the enigmatic damselfly Ischnura pumilio" / Journal of Insect Conservation, 2010. Volume 14. Number 6. pp. 689–700.
12. Allen K. "Science Report – The ecology and conservation of threatened damselflies" Science report: SC040027/SR1. 2009 - 142 pp.
13. Mauersberger R., Brauner O. "Kleine Pechlibelle – Ischnura pumilio (Charpentier 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 58-59.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
6. Smallshire D., Swash A. "Scarce Blue-tailed Damselfly. Ischnura pumilio" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 94-96.
7. Smallshire D., Swash A. "Small Bluetail. Ischnura pumilio" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 102-103.
8. Moreno-Benítez J. M., Ripoll J. "Ischnura pumilio" / in: Dragonflies along the Great Málaga Path and in the Province. Identification Guide, 2018. pp. 72-73.
9. Olberg S., Lønnve O. J. "Ischnura pumilio (Charpentier, 1825) (Odonoata, Coenagrionidae) in Norway" / Norwegian Journal of Entomology 59, 229–233. 2012.
10. Cham S., Nelson B., Parr A., Prentice S., Smallshire D., Taylor, P. "Atlas of the Dragonflies in Britain and Ireland". Centre for Ecology & Hydrology, 2014. - 288 pp.
11. Allen K. A., Le Duc M. G., Thompson D. J. "Habitat and conservation of the enigmatic damselfly Ischnura pumilio" / Journal of Insect Conservation, 2010. Volume 14. Number 6. pp. 689–700.
12. Allen K. "Science Report – The ecology and conservation of threatened damselflies" Science report: SC040027/SR1. 2009 - 142 pp.
13. Mauersberger R., Brauner O. "Kleine Pechlibelle – Ischnura pumilio (Charpentier 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 58-59.
