ЛЮТКА-НЕВЕСТА
Lestes sponsa Hansemann, 1823

♂ Фото © Виктор Рындевич, Налибокская пуща
Lestes sponsa Hansemann, 1823 (лютка-невеста)
Задняя часть головы целиком черная, желтых пятен нет. ♂ и ♀ окрашены одинаково. Грудь и брюшко сверху металлически-бронзово-зеленые, часто сильно блестящие. Окраска схожа с окраской Lestes dryas, но несколько светлее. У самцов с возрастом появляется бледно-голубой налет, который у половозрелых особей распространяется на дорсальную поверхность I и II сегментов брюшка и задние сегменты брюшка, а также на бока груди снизу. Самки более тускло-зеленые с участками розового или бежевого цвета по бокам груди и брюшка. Тело становится бронзовым у старых особей. Птеростигма узкая, ее ширина меньше 1/3 ее длины, длина в 2 ячейки. Птеростигма всех крыльев расположена одинаково далеко от вершины крыла, у молодых она светлая, затем темнеет и становится от коричневой до черноватой. В покое крылья держит полураскрытыми - промежуточное положение между большинством равнокрылых и разнокрылых стрекоз.
♂: Глаза голубые. Похож на L. dryas, но тело более тусклого цвета; срединный шов груди с тонкой, плечевые швы с более широкой желтоватой полоской; бока груди желтоватые, на втором шве тонкая черная полоска; у взрослых такой же беловато-голубой налет на переднеспинке и боках груди, I(II) и двух последних брюшных сегментах IX-X (иногда и VIII). Нижние анальные придатки прямые палочкообразные, на конце не расширены (иногда только слабо утолщены); на внутренней стороне верхних анальных придатков при основании такой же острый зубец, но второй зубец приближен к первому и одинаковой с ним величины, между ними более мелкие зубчики.
♀: Глаза коричневые. Брюшко отчетливо толще, чем у самца. Основной цвет более бронзовый; срединный шов с тонкой, плечевые швы с довольно широкой желтой полосой; бока груди желтые без черной полоски на втором шве. Боковые стороны переднегруди в основном светлые, только близ спинки с небольшим зеленым пятном. Брюшко без голубого цвета и с бежевым низом (хотя с возрастом изредка покрывается сливовым оттенком). Бронзовый значок на I тергите брюшка спереди более узкий с сильно срезанными передними углами. На сегменте II пара округлых темных пятен. Другие второстепенные особенности включают бледные нижние доли переднеспинки, изолированное металлическое пятно чуть выше основания средней ноги и тонкие желтоватые доплечевые полосы. Яйцеклад выступающий, едва достигает вершины сегмента X.
Генитальные заслоночки оканчиваются на уровне конца X членика брюшка. Верхние анальные придатки желтоватые с черной вершиной.
Неполовозрелые обоих полов металлически изумрудного цвета с теплыми охристыми нижними частями тела и созревают в течение 2 недель или около того, что намного дольше, чем у L. dryas.
При определении целесообразно осматривать анальные придатки самцов (проверить, являются нижние придатки почти прямыми или булавовидными, если смотреть сверху) и створки яйцеклада самок (проверить, не выступают ли они за вершину брюшного сегмента X), чтобы отделить этот вид от L. dryas.
Лет: конец мая - середина сентября.
Длина 35-39 мм, брюшко 25-30 мм, заднее крыло 18-22 мм. Встречаются карликовые формы самок с длиной тела 24-27 мм.
♂: Глаза голубые. Похож на L. dryas, но тело более тусклого цвета; срединный шов груди с тонкой, плечевые швы с более широкой желтоватой полоской; бока груди желтоватые, на втором шве тонкая черная полоска; у взрослых такой же беловато-голубой налет на переднеспинке и боках груди, I(II) и двух последних брюшных сегментах IX-X (иногда и VIII). Нижние анальные придатки прямые палочкообразные, на конце не расширены (иногда только слабо утолщены); на внутренней стороне верхних анальных придатков при основании такой же острый зубец, но второй зубец приближен к первому и одинаковой с ним величины, между ними более мелкие зубчики.
♀: Глаза коричневые. Брюшко отчетливо толще, чем у самца. Основной цвет более бронзовый; срединный шов с тонкой, плечевые швы с довольно широкой желтой полосой; бока груди желтые без черной полоски на втором шве. Боковые стороны переднегруди в основном светлые, только близ спинки с небольшим зеленым пятном. Брюшко без голубого цвета и с бежевым низом (хотя с возрастом изредка покрывается сливовым оттенком). Бронзовый значок на I тергите брюшка спереди более узкий с сильно срезанными передними углами. На сегменте II пара округлых темных пятен. Другие второстепенные особенности включают бледные нижние доли переднеспинки, изолированное металлическое пятно чуть выше основания средней ноги и тонкие желтоватые доплечевые полосы. Яйцеклад выступающий, едва достигает вершины сегмента X.
Генитальные заслоночки оканчиваются на уровне конца X членика брюшка. Верхние анальные придатки желтоватые с черной вершиной.
Неполовозрелые обоих полов металлически изумрудного цвета с теплыми охристыми нижними частями тела и созревают в течение 2 недель или около того, что намного дольше, чем у L. dryas.
При определении целесообразно осматривать анальные придатки самцов (проверить, являются нижние придатки почти прямыми или булавовидными, если смотреть сверху) и створки яйцеклада самок (проверить, не выступают ли они за вершину брюшного сегмента X), чтобы отделить этот вид от L. dryas.
Лет: конец мая - середина сентября.
Длина 35-39 мм, брюшко 25-30 мм, заднее крыло 18-22 мм. Встречаются карликовые формы самок с длиной тела 24-27 мм.

♀ Фото © Виталий Сирота, г. Минск
Населяет самые разнообразные стоячие и текущие водоемы, особенно с кислой реакцией воды, богатые водной растительностью и хорошо развитым поясом тростников. Большинство местообитаний относятся к небольшим хорошо освещенным водоемам со стоячей водой. На озерах встречается сравнительно редко, где занимает обширные и в то же время почти свободные от рыбы зоны заиления, а собственно само озеро избегается. Летает довольно слабо и предпочитает высокую, густую растительность, в которой сидит довольно незаметно. Взрослые особи держатся возле стоячих водоемов, медленно летая над травой и среди ветвей прибрежных кустарников. В отличие от некоторых других видов равнокрылых стрекоз, иногда устраивается на ночлег у кромки воды. В популяциях самцы численно преобладают над самками. Они зимуют.
Размножается в очень широком диапазоне местообитаний от эвтрофных канав до болотистых водоемов. Предпочитает мелководье с обилием высокой надводной и маргинальной растительности, такой как тростник, осока или хвощ, хотя было зарегистрировано размножение в прудах, лишенных как подводных, так и надводных растений, если присутствует подходящая растительность на берегу водоемов.
Размножается в очень широком диапазоне местообитаний от эвтрофных канав до болотистых водоемов. Предпочитает мелководье с обилием высокой надводной и маргинальной растительности, такой как тростник, осока или хвощ, хотя было зарегистрировано размножение в прудах, лишенных как подводных, так и надводных растений, если присутствует подходящая растительность на берегу водоемов.

♂ Фото © Владимир Бондарь, Могилевский р-н
Самцы территориальны, например, в Британии их плотность редко превышает 110 особей на 100 м уреза воды. Совокупление происходит недалеко или возле воды и имеет длительную продолжительность (28-69 минут).
Яйца откладываются в ткани (стебли, листья) водных растений (Iris, Heleocharis, Juncus, Scirpus, Alisma, Sparganium и мн. др.). Самки, обычно сопровождаемые самцами, откладывают яйца или в стебли растений, находящихся под водой, для чего они вместе погружаются в воду и постепенно опускаясь вниз, самка откладывает яйца, или же на растения, оказавшиеся временно вне воды из-за совершенного высыхания водоема. Самки могут полностью погружаться в воду, иногда оставаясь под ней до 30 минут.
Яйца удлиненные, располагаются в 1 или 2 вертикальных ряда, содержащих около 20 яиц: расстояние между яйцами всегда постоянно, приблизительно 0.5 или 0.75 см. Яйца этого вида, отложенные поздней осенью, устойчивы к высыханию и низким температурам, перезимовывают и предличинки вылупляются только весной следующего года при новом заполнении водоема водой и при благоприятных термических условиях. Предличинка при выходе из яйца делает толчкообразное движение и выпадает на поверхность воды, затем ее покрытие лопается и через несколько минут наружу выходит уже личинка.
Яйца откладываются в ткани (стебли, листья) водных растений (Iris, Heleocharis, Juncus, Scirpus, Alisma, Sparganium и мн. др.). Самки, обычно сопровождаемые самцами, откладывают яйца или в стебли растений, находящихся под водой, для чего они вместе погружаются в воду и постепенно опускаясь вниз, самка откладывает яйца, или же на растения, оказавшиеся временно вне воды из-за совершенного высыхания водоема. Самки могут полностью погружаться в воду, иногда оставаясь под ней до 30 минут.
Яйца удлиненные, располагаются в 1 или 2 вертикальных ряда, содержащих около 20 яиц: расстояние между яйцами всегда постоянно, приблизительно 0.5 или 0.75 см. Яйца этого вида, отложенные поздней осенью, устойчивы к высыханию и низким температурам, перезимовывают и предличинки вылупляются только весной следующего года при новом заполнении водоема водой и при благоприятных термических условиях. Предличинка при выходе из яйца делает толчкообразное движение и выпадает на поверхность воды, затем ее покрытие лопается и через несколько минут наружу выходит уже личинка.

♀ Фото © Владимир Бондарь, Могилевский р-н
Данные об откладке яиц у этого вида разнятся регионально. Например, в Швейцарии отмечены нередкие случаи, когда самки откладывали яйца намного выше уровня воды (на 50 см выше), а целом самки откладывают яйца начиная чуть выше уровня воды и затем немного погружаясь в воду. В Чехии также считается, что вид редко откладывает яйца под водой и только в подходящих условиях. К тому же, количество отложенных яиц отличается от данных Поповой - в бороздку от 2 до 4 яиц. Учитывая, что данные Поповой, хотя и относятся к территории бывшего СССР, но датированы до 1950-х гг., а данные из южной Чехии относятся к 2010 и 2011 гг. С другой стороны, климатические условия в Чехии отличаются от белорусских и об этом надо помнить.
Согласно чешским специалистам, откладка яиц происходит сразу после копуляции и обычно пока самцы остаются вместе с самками в тандеме. В исключительных случаях самки откладывают яйца в одиночку. Вид в Чехии обычно откладывает свои яйца в стебли надводных растений (например, Juncus, Equisetum, Schoenoplectus или Eleocharis spp.), а также в ткани листьев галофитов, например, Iris или Stratiotes spp. Самка сначала проникает яйцекладом в растительную ткань, а затем откладывает в бороздку от 2 до 4 яиц. Кладки ориентированы по линии вдоль волокон растительной ткани.
Частота подводной яйцекладки на различных участках изучаемого региона была относительно низкой (менее 20%), но пропорциональное использование этой стратегии в отдельных местах было высоким. Подводная яйцекладка наблюдалась только на трех из 16 участков. Во всех этих трех местах зарегистрировано большее количество яиц, отложенных под водой, чем над ее поверхностью. Всего подсчитано 7699 яиц, из них 4759 яиц (62%) было отложено под водой.
Согласно чешским специалистам, откладка яиц происходит сразу после копуляции и обычно пока самцы остаются вместе с самками в тандеме. В исключительных случаях самки откладывают яйца в одиночку. Вид в Чехии обычно откладывает свои яйца в стебли надводных растений (например, Juncus, Equisetum, Schoenoplectus или Eleocharis spp.), а также в ткани листьев галофитов, например, Iris или Stratiotes spp. Самка сначала проникает яйцекладом в растительную ткань, а затем откладывает в бороздку от 2 до 4 яиц. Кладки ориентированы по линии вдоль волокон растительной ткани.
Частота подводной яйцекладки на различных участках изучаемого региона была относительно низкой (менее 20%), но пропорциональное использование этой стратегии в отдельных местах было высоким. Подводная яйцекладка наблюдалась только на трех из 16 участков. Во всех этих трех местах зарегистрировано большее количество яиц, отложенных под водой, чем над ее поверхностью. Всего подсчитано 7699 яиц, из них 4759 яиц (62%) было отложено под водой.

♂ Фото © Владимир Бондарь, Могилевский р-н
В Чехии общими признаками местообитаний, в которых пары откладывали яйца под водой, были прозрачность воды и относительно низкая степень затенения. Оказалось, что, хотя ни один конкретный вид растений не был в значительной степени благоприятен для подводной яйцекладки, общая морфологическая структура растения (особенно диаметр и прочность стебля) была важна. Самки явно предпочитали тонкие и мягкие стебли Eleocharis, Juncus и Schoenoplectus стеблям Equisetum или Typha для подводной откладки яиц. Однако не все из этих растений были обычными на всех участках.
Подводная яйцекладка происходила преимущественно в ясные теплые дни в периоды, когда температура воздуха превышала 22°С. Во всех наблюдениях самки откладывали яйца тандемно. Оба партнера приземлялись на всплывшие части растений. Вскоре после посадки самка изгибала брюшко и начинала ощупывать субстрат яйцекладом. Во всех случаях самка начинала кладку над поверхностью воды и непрерывно откладывала яйца, двигаясь задом под воду. Яйцекладка ненадолго прерывалась только тогда, когда оба партнера складывали крылья и появлялись над поверхностью воды. Если тандем нарушался, обе особи тут же пытались выбраться из воды. Самцы во всех случаях оставались связанными с самками на протяжении всей подводной яйцекладки. Яйцекладущие пары в среднем проводили под поверхностью воды 338 секунд, максимальная продолжительность пребывания под водой составила 1268 секунд. Также наблюдались пары, которые, вероятно, не откладывали всю кладку за одно погружение и неоднократно откладывали яйца под водой, причем один тандем делал это 7 раз в течение нескольких минут.
Максимальная зарегистрированная глубина откладки яиц составила 55 см от поверхности воды, однако она часто приходилась на дно водоема. Большинство наблюдаемых яйцекладок находилось на максимальных глубинах в диапазоне 10-20 см.
Подводная яйцекладка происходила преимущественно в ясные теплые дни в периоды, когда температура воздуха превышала 22°С. Во всех наблюдениях самки откладывали яйца тандемно. Оба партнера приземлялись на всплывшие части растений. Вскоре после посадки самка изгибала брюшко и начинала ощупывать субстрат яйцекладом. Во всех случаях самка начинала кладку над поверхностью воды и непрерывно откладывала яйца, двигаясь задом под воду. Яйцекладка ненадолго прерывалась только тогда, когда оба партнера складывали крылья и появлялись над поверхностью воды. Если тандем нарушался, обе особи тут же пытались выбраться из воды. Самцы во всех случаях оставались связанными с самками на протяжении всей подводной яйцекладки. Яйцекладущие пары в среднем проводили под поверхностью воды 338 секунд, максимальная продолжительность пребывания под водой составила 1268 секунд. Также наблюдались пары, которые, вероятно, не откладывали всю кладку за одно погружение и неоднократно откладывали яйца под водой, причем один тандем делал это 7 раз в течение нескольких минут.
Максимальная зарегистрированная глубина откладки яиц составила 55 см от поверхности воды, однако она часто приходилась на дно водоема. Большинство наблюдаемых яйцекладок находилось на максимальных глубинах в диапазоне 10-20 см.

♂ Фото © Владимир Бондарь, Могилевский р-н
Нимфа. Тело длинное, стройное, окраска варьирует от светлой красновато-коричневой до темной серовато-коричневой; у более темных зкземпляров вентральная сторона также сероватая. Чаще встречаются темные особи. Длина, включая жаберные пластинки, 25,5-26,5 мм, ширина 2,25 мм. Маска длинная, узкая, в дистальной части расширенная наподобие ложки, в состоянии покоя заходит за середину основания задней пары ног; внешний зубец дистального края боковой лопасти маски сильно изогнутый, намного больше внутреннего, между ними расположены 7 маленьких зубчиков. Переднегрудь дорсально четырехугольная, с более широкой задней стороной; среднегрудные дыхальца темные, ясно видимые. Крыловые чехлы короткие, едва достигают IV сегмента, длиной 5,5 мм. Бедра и голени с очень тонкими темными полосками, из которых более широкая темная проходит по внешней стороне бедер; на бедрах несколько дистальных темных колец, а на голенях одно очень узкое дистальное темное кольцо: дистальные концы бедер, голеней и члеников лапок темные; длина передних ног 7,5 мм, средних 9,5 мм и задних 12 мм. Брюшко с дорсальной стороны с беловатой узкой продольной центральной полоской, окаймленной весьма узкой расплывчатой темной полосой; за этой полосой следует с каждой стороны по 1 широкой светлой и 1 еще более широкой темной полосе, в которую на конце каждого сегмента врезается более светлая полоса, занимающая 2/3 длины каждого сегмента; на вентральной стороне, за светлым боковым краем от 1 до начала IX сегмента, проходит темная продольная линия; IX и Х сегменты посредине с дорсальной стороны приподняты в виде гребня (киля); латеральные шипы на V-IX сегментах маленькие, прямые, четкие, очень острые. Хвостовые жабры в виде листообразных пластинок, очень длинных, равных 2/3 длины всего брюшка, на конце округлые, с небольшим острием, длиной почти 10 мм; форма и окраска их варьирует от светло-золотисто-коричневой до почти черной; края пластинок снабжены короткими шипами. Створки яйцеклада самки длинные, боковые створки достигают базальной трети Х сегмента; стили, верхние и средние створки одинаковой длины, доходят до середины основной части жабер.

♂ juv Фото © Владимир Бондарь, Могилевский р-н
Типичный лимнофил, развивающийся в небольших стоячих водоемах: озерках, прудах, болотах, канавах, заводях, лужах, в том числе и в пересыхающих. Предпочитает мелкие (глубиной до 1-1,3 м), хорошо прогреваемые участки с богатой водной растительностью. Может встречаться также и на мелководье медленно текущих рек и ручьев. Личинки L. sponsa, как и личинки других видов рода, характерные фитофилы: заросли водных растений являются непременным условием их жизни в тех или иных водоемах. Ведет донный и придонный образ жизни, ползая и устраивая засады на дне или в нижней части стебля водных растений. Пищей личинок являются главным образом низшие ракообразные (Cladocera и Copepoda). Из Cladocera преобладают формы мелких заросших водоемов: Acroperus harpea, Ceriodaphnia reticulata, Scapholeberis mucronata, Chydorus sphaericus, Sida crystallina, а также некоторые Bosmina и другие обычные планктонные виды. Известно, что многие из перечисленных форм Cladocera в яркие солнечные дни держатся у дна. Следовательно, и прожорливым личинкам, каковыми являются личинки рода Lestes, приходится днем спускаться иногда на дно водоемов, где они и лежат спокойно, выжидая свою добычу. Это предположение подтверждается наблюдениями, когда находили личинок этого вида как между водными растениями, на глубине 0,1-1,0 м, так и на дне (в меньшем количестве).
Появление личинок происходит в конце апреля - начале мая. Растут они очень быстро и, перелиняв до 10-11 раз, заканчивают свое развитие за 2,5-3 месяца, хотя личинки последнего возраста иногда зимуют. Таким образом, этот вид может переживать высыхание среды обитания во время засухи в конце лета. Цикл развития одногодичный. Однако имаго достигают зрелости необычно долго (16-30 дней). Имаго появляются на растительности на высоте около 50 см над водой. Вид можно обнаружить почти со всеми другими видами, которые размножаются в стоячей воде.
Появление личинок происходит в конце апреля - начале мая. Растут они очень быстро и, перелиняв до 10-11 раз, заканчивают свое развитие за 2,5-3 месяца, хотя личинки последнего возраста иногда зимуют. Таким образом, этот вид может переживать высыхание среды обитания во время засухи в конце лета. Цикл развития одногодичный. Однако имаго достигают зрелости необычно долго (16-30 дней). Имаго появляются на растительности на высоте около 50 см над водой. Вид можно обнаружить почти со всеми другими видами, которые размножаются в стоячей воде.

♀, поедающая Enallagma cyathigerum. Фото © Виктор Рындевич

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
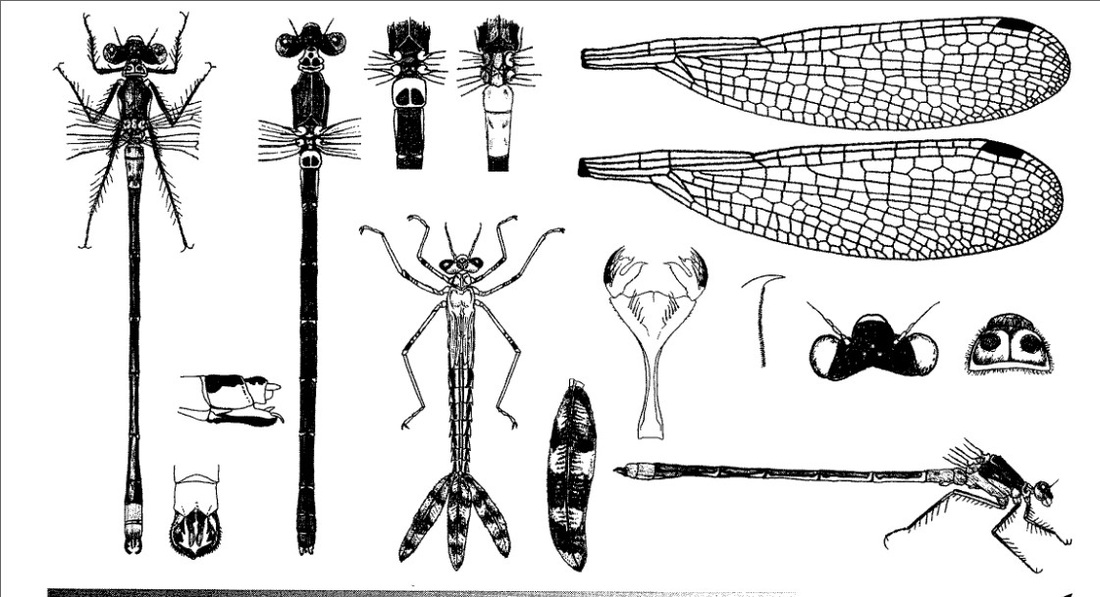
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Dolný A., Helebrandová J., Rusková T., Šigut M., Harabiš F. "Ecological aspects of underwater oviposition in Lestes sponsa (Odonata: Lestidae)" / Odonatologica 43(3/4) 2014: 183-197.
9. Monnerat C. "Lestes sponsa (Hansemann, 1823) Leste fiancé - Gemeinde Binsenjungfer"/ in: Wildennuth H., Gonseth Y., Maibach A. (éds.) Odonata. Les Libellules de Suisse. Fauna Helvetica 11. Centre suisse de cartographie de la faune Schvveizerische Entornologische Gesellschaft, 2005. pp. 96-99.
10. Smallshire D., Swash A. "Emerald Damselfly. Lestes sponsa" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 64-65.
11. Smallshire D., Swash A. "Common Spreadwing. Lestes sponsa" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 20-21.
12. Brauner O., Petzold F. "Gemeine Binsenjungfer – Lestes sponsa (Hansemann 1823)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 22-23.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Dolný A., Helebrandová J., Rusková T., Šigut M., Harabiš F. "Ecological aspects of underwater oviposition in Lestes sponsa (Odonata: Lestidae)" / Odonatologica 43(3/4) 2014: 183-197.
9. Monnerat C. "Lestes sponsa (Hansemann, 1823) Leste fiancé - Gemeinde Binsenjungfer"/ in: Wildennuth H., Gonseth Y., Maibach A. (éds.) Odonata. Les Libellules de Suisse. Fauna Helvetica 11. Centre suisse de cartographie de la faune Schvveizerische Entornologische Gesellschaft, 2005. pp. 96-99.
10. Smallshire D., Swash A. "Emerald Damselfly. Lestes sponsa" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 64-65.
11. Smallshire D., Swash A. "Common Spreadwing. Lestes sponsa" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 20-21.
12. Brauner O., Petzold F. "Gemeine Binsenjungfer – Lestes sponsa (Hansemann 1823)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 22-23.
