НЕХАЛЕНИЯ КРАСИВАЯ
Nehalennia speciosa Charpentier, 1840

Фото © Денис Китель, Малоритский р-н (Брестская обл.)
Nehalennia speciosa Charpentier, 1840 (нехаления красивая)
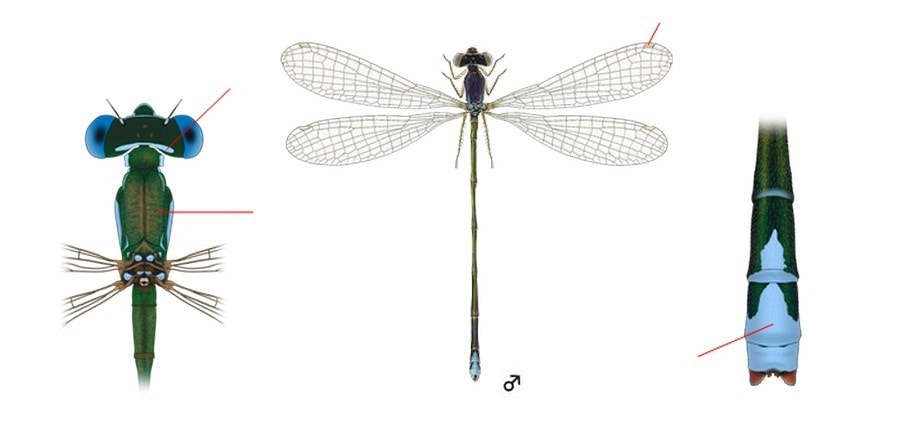
Самая маленькая стрекоза Беларуси, как и всей материковой Европы. Голова широкая, сверху вдвое шире своей длины. Лоб с острым поперечным краем спереди от антенн, в профиль выглядит заостренным. Затылок сверху темный, с узкой светлой дуговидной поперечной полосой, обычно голубой у обоих полов, но иногда желтоватой у самок. Затылочные пятна отсутствуют. Глаза разного цвета в зависимости от возраста и пола: голубые, светло-зеленые, коричневые или двухцветные с темным верхом. Верхняя часть тела металлически блестящая, ярко-зеленая (с возрастом бронзовая). Бока груди, а также ее низ и нижняя часть брюшка светло-голубые. Крылья прозрачные, с очень разреженным жилкованием, в покое держит сложенными. Крылья в покое не заходят на V сегмент брюшка (2/3 длины брюшка). Птеростигма узкая (равна 1 ячейке), светлая. Ноги черные или темно-серые.
♂: Глаза светло-голубые. Полоска позади глаз, бока груди, X членик брюшка целиком и VIII-IX частично голубые, сужающиеся в форме «клина». Кончик сегмента X имеет на вершине пару крошечных шипов. Верхние придатки относительно большие и тупые. Низ груди и ноги светлые; последние снаружи буроватые. Птеростигма желтая.
♀: Окрашена как самец, но с возрастом светло-голубые участки сменяется от оранжевых до темно-коричневых. Птеростигма белая. Ноги желтоватые.
Существует вероятность спутать с лютками (Lestes spp.) из-за металлически-зеленой окраски и голубого «хвоста». Однако виды Lestes намного крупнее и когда сидят, держат крылья полураскрытыми. Старые самки оранжевого цвета внешне напоминают самок Ischnura elegans, но металлический зеленый цвет и «хвостовой фонарь» у последних отсутствуют.
Длина 24-26 мм, брюшко 19-22 мм, заднее крыло 11-16 мм.
Лет: начало июня - конец августа.
♂: Глаза светло-голубые. Полоска позади глаз, бока груди, X членик брюшка целиком и VIII-IX частично голубые, сужающиеся в форме «клина». Кончик сегмента X имеет на вершине пару крошечных шипов. Верхние придатки относительно большие и тупые. Низ груди и ноги светлые; последние снаружи буроватые. Птеростигма желтая.
♀: Окрашена как самец, но с возрастом светло-голубые участки сменяется от оранжевых до темно-коричневых. Птеростигма белая. Ноги желтоватые.
Существует вероятность спутать с лютками (Lestes spp.) из-за металлически-зеленой окраски и голубого «хвоста». Однако виды Lestes намного крупнее и когда сидят, держат крылья полураскрытыми. Старые самки оранжевого цвета внешне напоминают самок Ischnura elegans, но металлический зеленый цвет и «хвостовой фонарь» у последних отсутствуют.
Длина 24-26 мм, брюшко 19-22 мм, заднее крыло 11-16 мм.
Лет: начало июня - конец августа.

♂ Фото © Денис Китель, Малоритский р-н (Брестская обл.)
Миниатюрная, скрытная стрекоза является очень локализованным обитателем болот и озер. В настоящее время встречается в основном в Восточной Европе (до Эстонии, Латвии, Литвы, северной и восточной Польши, северной и южной Беларуси и, возможно, 52–54° с.ш. в России) и в Баварском предальпийском регионе. Во всех остальных районах вид сохранился в единичных или редких локальных популяциях.
Ареал заметно сократился с 1960-х годов из-за факторов окружающей среды. Один из шести видов этого рода, остальные пять обитают в Америке.
В Беларуси вид отмечен в основном Витебской, Гомельской и Брестской, а также в Минской областях, однако в последней его распространение почти неизвестно. Предполагается, что обнаружен лишь небольшой процент местонахождений нехалении в Беларуси. Вся белорусская популяция, скорее всего, еще многочисленна и находится в хорошем состоянии. Два местонахождения находятся в озерных районах северо-востока Беларуси, еще четыре — на юге, в заболоченной области Полесья. И северный и южный участки, безусловно, являются центрами распространения вида в стране. Особого внимания заслуживает крупная текущая популяция на болоте Симоничи в Полесье, возможно, вид не менее обилен в заказнике «Споровский».
Ареал заметно сократился с 1960-х годов из-за факторов окружающей среды. Один из шести видов этого рода, остальные пять обитают в Америке.
В Беларуси вид отмечен в основном Витебской, Гомельской и Брестской, а также в Минской областях, однако в последней его распространение почти неизвестно. Предполагается, что обнаружен лишь небольшой процент местонахождений нехалении в Беларуси. Вся белорусская популяция, скорее всего, еще многочисленна и находится в хорошем состоянии. Два местонахождения находятся в озерных районах северо-востока Беларуси, еще четыре — на юге, в заболоченной области Полесья. И северный и южный участки, безусловно, являются центрами распространения вида в стране. Особого внимания заслуживает крупная текущая популяция на болоте Симоничи в Полесье, возможно, вид не менее обилен в заказнике «Споровский».

♀ Фото © Семён Левый, Малоритский р-н, Брестская обл.
К среде обитания очень специфические требования, размножение в мелких кислых водоемах торфяных болот или окраин озер. Они должны быть хорошо защищены зарослями осоки (Carex spp.) или, в исключительных случаях, другими видами, таких как молиния (Molinia caerulea) или приречный хвощ (Equisetum fluviatile). Carex lasiocarpa - наиболее частый и многочисленный представитель узколистных растений, предпочитаемых видом. C. limosa, хотя обычно встречается немного реже и менее многочислен, также всегда выбирается нехаленией. Обычная и многочисленная C. rostrata также может использоваться видом, но если встречается C. lasiocarpa и C. limosa, то они являются явно предпочтительными. Кроме того, в двух местах в Латвии C. elata был ведущим видом в среде обитания нехалении. Помимо других типичных компонентов местообитаний, т.е. Sphagnum sp., Utricularia sp., Menyanthes trifoliata, Comarum palustre, Lysimachia thyrsiflora, Equisetum limosum, в Латвии встречаются довольно нетипичные виды растений, такие как Thelypteris palustris, Typha latifolia и Hydrocharis morsus-ranae.
В Швейцарии населяет только заливные осоковые луга и покрытые осокой канавы промежуточных болот. Глубина водоема составляет в среднем около 10 см и может варьироваться от 3 до 30 см, но он никогда не должен полностью пересыхать. Вода с низким содержанием карбонатов и других электролитов и от слабой до умеренно кислой. Растительность в основном состоит из узколистных видов осок, таких как Carex limosa, C. lasiocarpa и C. elata, со средней высотой 35-50 см. К ним могут примешиваться широколиственные цветковые растения. Общее покрытие составляет не более 70%. Под водой растительность состоит из мхов и видов пузырчатки Utricularia. Хорошими указателями на наличие нехалении являются U. intermedia и U. minor.
В Швейцарии населяет только заливные осоковые луга и покрытые осокой канавы промежуточных болот. Глубина водоема составляет в среднем около 10 см и может варьироваться от 3 до 30 см, но он никогда не должен полностью пересыхать. Вода с низким содержанием карбонатов и других электролитов и от слабой до умеренно кислой. Растительность в основном состоит из узколистных видов осок, таких как Carex limosa, C. lasiocarpa и C. elata, со средней высотой 35-50 см. К ним могут примешиваться широколиственные цветковые растения. Общее покрытие составляет не более 70%. Под водой растительность состоит из мхов и видов пузырчатки Utricularia. Хорошими указателями на наличие нехалении являются U. intermedia и U. minor.

♂ Фото © Семён Левый, Малоритский р-н, Брестская обл.
В Европе участки, заселенные видом, включают как первичные, так и вторичные местообитания, при этом предпочтение отдается первым. В то время как в центре ареала вид почти полностью ограничен первичными местообитаниями, в приграничных районах, очевидно, из-за обширного их разрушения, доля колонизированных вторичных местообитаний значительно выше. Например, за исключением одного местонахождения в Литве известны только первичные местообитания, а в Польше обнаружено только 4 вторичных местообитания, в отличие от не менее 40 первичных местообитаний. С другой стороны, в Швейцарии и Нижней Саксонии этот вид был зарегистрирован исключительно из вторичных местообитаний. В Баварии и Баден-Вюртемберге встречается в обоих типах местообитаний.
В Швейцарии вид известен только из вторичных биотопов. Это бывшие зарастающие торфяные выработки или канавы на осоковых болотах, которые питаются за счет грунтовых вод. Там, где популяции нехалении все еще существуют сегодня, растительный покров находится поверх слоя воды или жидкого торфяного ила толщиной не менее 2 м. Популяции, по-видимому, могут выжить только там, где ковер растительности остается на большой площади после опускания уровня воды в засушливые периоды. После достаточного количества осадков растительный покров вновь поднимается вместе с подъемом уровня грунтовых вод. Температура воды может сильно колебаться, но в среднем выше, чем слое воздуха над водой.
В Швейцарии вид известен только из вторичных биотопов. Это бывшие зарастающие торфяные выработки или канавы на осоковых болотах, которые питаются за счет грунтовых вод. Там, где популяции нехалении все еще существуют сегодня, растительный покров находится поверх слоя воды или жидкого торфяного ила толщиной не менее 2 м. Популяции, по-видимому, могут выжить только там, где ковер растительности остается на большой площади после опускания уровня воды в засушливые периоды. После достаточного количества осадков растительный покров вновь поднимается вместе с подъемом уровня грунтовых вод. Температура воды может сильно колебаться, но в среднем выше, чем слое воздуха над водой.

© Семён Левый, Малоритский р-н, Брестская обл.
В восточной Европе обитает почти исключительно на верховых болотах или вдоль мелководных и заболоченных окраин ледниковых озер со сплавинами из сфагнума, поросшими средней густоты зарослями узколистных осок.
Хотя подчеркивалось, что вид предпочитает небольшие водоемы (до 1 га) в Польше и странах Балтии, озера, населяемые этим видом в Латвии, часто превышают 1 га. 14 из 34 озер (41%) имеют размер от 1 до 10 га, а еще 9 озер (26%) превышают 10 га, 4 из них даже превышают 1000 га. Это показывает, что вид не избегает более крупных, казалось бы, неподходящих водоемов и может заселять небольшие подходящие участки в пределах заведомо неподходящих местообитаний. На некоторых средних и крупных озерах N. speciosa может использовать растительность, не граничащую непосредственно с зеркалом воды, а входящую в состав сопутствующих комплексов болот. Таким образом, такие более крупные озера, особенно дистрофные, мезотрофные и слабоэвтрофные с богатой и обильной окружающей растительностью, не следует исключать априори.
Хотя подчеркивалось, что вид предпочитает небольшие водоемы (до 1 га) в Польше и странах Балтии, озера, населяемые этим видом в Латвии, часто превышают 1 га. 14 из 34 озер (41%) имеют размер от 1 до 10 га, а еще 9 озер (26%) превышают 10 га, 4 из них даже превышают 1000 га. Это показывает, что вид не избегает более крупных, казалось бы, неподходящих водоемов и может заселять небольшие подходящие участки в пределах заведомо неподходящих местообитаний. На некоторых средних и крупных озерах N. speciosa может использовать растительность, не граничащую непосредственно с зеркалом воды, а входящую в состав сопутствующих комплексов болот. Таким образом, такие более крупные озера, особенно дистрофные, мезотрофные и слабоэвтрофные с богатой и обильной окружающей растительностью, не следует исключать априори.

♂ Фото © Денис Китель, Россонский р-н, Витебская обл.
Среду обитания можно разделить на четыре основных типа: 1) озера; 2) болота как первичные среды обитания; 3) топи и болота как первичные и вторичные среды обитания, 4) искусственные водоемы и пруды как вторичные места обитания. Озера, населенные этим видом, в основном небольшие, в Польше и Прибалтике обычно имеют длину 50-100 м и ширину 40-100 м и, по крайней мере, частично окаймлены узкой полосой специфической низкой переходно-болотной растительности. Что касается сфагновых болот и топей, то они либо неглубоко затоплены, либо, что чаще, представляют собой несколько небольших луж и мелководных котловин.
Вторичные местообитания представлены влажными подстилочными лугами, зарегистрированными в Баварии, некоторыми болотами, например, в пойме Нежарки в Чехии, двумя бывшими рыбными прудами в Люксембурге и Баден-Вюртемберге, но в основном в затопленных бывших карьрах. Заброшенные торфяные выработки, частично заросшие, являются основным типом вторичных местообитаний в Польше, Бранденбурге, Нижней Саксонии, Шлезвиг-Гольштейне, Баварии и Швейцарии. Единственным исключением являются до сих пор используемые глиняные карьеры в польской Верхней Силезии, заселенные этим видом только в самых старых частях и густо заросшие осокой.
Как небольшие изолированные объекты, встречающиеся в основном в Польше и Прибалтике, так и реже фрагменты крупных комплексов болотных угодий, как, например, на Споровском болоте в Беларуси и в региональном парке Гирутишкис в Литве являются убежищем нехалении.
Вторичные местообитания представлены влажными подстилочными лугами, зарегистрированными в Баварии, некоторыми болотами, например, в пойме Нежарки в Чехии, двумя бывшими рыбными прудами в Люксембурге и Баден-Вюртемберге, но в основном в затопленных бывших карьрах. Заброшенные торфяные выработки, частично заросшие, являются основным типом вторичных местообитаний в Польше, Бранденбурге, Нижней Саксонии, Шлезвиг-Гольштейне, Баварии и Швейцарии. Единственным исключением являются до сих пор используемые глиняные карьеры в польской Верхней Силезии, заселенные этим видом только в самых старых частях и густо заросшие осокой.
Как небольшие изолированные объекты, встречающиеся в основном в Польше и Прибалтике, так и реже фрагменты крупных комплексов болотных угодий, как, например, на Споровском болоте в Беларуси и в региональном парке Гирутишкис в Литве являются убежищем нехалении.

♀ Фото © Александр Месяц, Борисовский р-н, Минская обл.
Между распространением вида и территориями, покрытыми плейстоценовыми ледниками, наблюдается поразительное совпадение, хотя некоторые местообитания находятся за пределами этой области. В северо-восточной Германии, северной Польше и странах Балтии места обитания в послеледниковых впадинах или неглубоких каналах часто колонизируются нехаленией. Такие места обитания представляют собой сфагновые болота с центральным небольшим озером или несколькими водоемами. В Швейцарии и на юго-востоке Германии распространение вида также в основном приурочено к ареалам, образованным плейстоценовым оледенением. Местностей, расположенных на больших равнинных территориях, меньшинство, типичным примером является болото Симоничи в заболоченном бассейне р. Припять на юге Беларуси.
В ядре ареала вида почти все местообитания находятся в лесистой местности. То же самое можно сказать и о многих локациях юго-западного пограничья. В Польше и Прибалтике вид чаще всего встречается в крупных лесных массивах и почти никогда на их опушках. На минеральных почвах это в большинстве случаев хвойные леса или, по крайней мере, в них преобладают хвойные компоненты, такие как сосна (Pinus sylvestris) и ель (Picea abies). В виде исключения небольшая изолированная местность близ Пороста (Западно-Поморское воеводство, Польша) окружена старым лиственным лесом с преобладанием бука (Fagus sylvatica). На северо-западе Польши, где ель почти отсутствует, участки N. speciosa окружены сосновыми насаждениями. В Литве, где эти два вида деревьев являются аборигенными, нехаления встречается в основном в сосновых лесах. Большинство болотных угодий в еловых лесах оказываются неподходящими для вида. Во многих случаях местообитания нехалении непосредственно окаймлены узкими древостоями на влажных торфяных грунтах, преимущественно сосновыми, реже березовыми (Betula pubescens).
В ядре ареала вида почти все местообитания находятся в лесистой местности. То же самое можно сказать и о многих локациях юго-западного пограничья. В Польше и Прибалтике вид чаще всего встречается в крупных лесных массивах и почти никогда на их опушках. На минеральных почвах это в большинстве случаев хвойные леса или, по крайней мере, в них преобладают хвойные компоненты, такие как сосна (Pinus sylvestris) и ель (Picea abies). В виде исключения небольшая изолированная местность близ Пороста (Западно-Поморское воеводство, Польша) окружена старым лиственным лесом с преобладанием бука (Fagus sylvatica). На северо-западе Польши, где ель почти отсутствует, участки N. speciosa окружены сосновыми насаждениями. В Литве, где эти два вида деревьев являются аборигенными, нехаления встречается в основном в сосновых лесах. Большинство болотных угодий в еловых лесах оказываются неподходящими для вида. Во многих случаях местообитания нехалении непосредственно окаймлены узкими древостоями на влажных торфяных грунтах, преимущественно сосновыми, реже березовыми (Betula pubescens).
Имаго и личинки, как правило, используют одни и те же местообитания, т. е. довольно узкую зону плавающих или затопленных слоев переплетенной растительности, граничащих с открытой водой озер и заводей, либо мелководные водоемы в болотах и топях, в сфагновых болотах это чаще всего небольшие понижения, в исключительных случаях водоемы внешней болотной зоны. В зависимости от температуры и влажности они живут в разных ярусах растительности.
В Польше и странах Балтии преобладают колонизированные местообитания, примыкающие к озерам, а на юго-западе Германии преобладают удаленные от крупных водоемов.
Взрослые особи покидают места размножения лишь в ограниченных масштабах. В Нижней Саксонии и Юго-Восточной Польше они были обнаружены среди растительности Molinia, прилегающей к бывшим торфяным выработкам. В Северной Польше и Прибалтике очень немного имаго было отмечено на сфагновом слое в нескольких метрах от воды, и только в 3 из примерно 40 исследованных местонахождений имаго встречались регулярно, но очень локально. Это говорит о том, что пропитанные водой слои сфагнума могут локально создать подходящие условия для личинок. Пока их развитие в сфагновой растительности подтверждено только на болоте Симоничи в Беларуси, где вся популяция N. speciosa была обнаружена вдали от открытой воды.
В Польше и странах Балтии преобладают колонизированные местообитания, примыкающие к озерам, а на юго-западе Германии преобладают удаленные от крупных водоемов.
Взрослые особи покидают места размножения лишь в ограниченных масштабах. В Нижней Саксонии и Юго-Восточной Польше они были обнаружены среди растительности Molinia, прилегающей к бывшим торфяным выработкам. В Северной Польше и Прибалтике очень немного имаго было отмечено на сфагновом слое в нескольких метрах от воды, и только в 3 из примерно 40 исследованных местонахождений имаго встречались регулярно, но очень локально. Это говорит о том, что пропитанные водой слои сфагнума могут локально создать подходящие условия для личинок. Пока их развитие в сфагновой растительности подтверждено только на болоте Симоничи в Беларуси, где вся популяция N. speciosa была обнаружена вдали от открытой воды.
Как правило, вид связан с определенной растительностью, но решающим фактором является ее текстура. Надводная растительность, населенная имаго, включает слой гелофитов над поверхностью воды и над непосредственно прилегающими слоями сфагнума. Несмотря на широкий спектр видов растений и некоторую географическую и экологическую изменчивость видового состава, Carex limosa и C. lasiocarpa, безусловно, являются наиболее важными компонентами, по крайней мере, в Европе. Один из этих видов, но чаще оба, отмечены на участках всего европейского ареала нехалении. В Польше, Литве, Беларуси, Швеции и Дании один или оба этих вида преобладают в местообитаниях, в то время как в других странах и регионах они, по крайней мере, составляют важный компонент спектра растений.
Подводная растительность в основном состоит из затопленных частей гелофитов, как живых, так и мертвых, и плавающих сфагновых и других мхов, таких как напр. Scorpidium scorpioides, Warnstorfia fluitans, Limprichtia revolvens, Calliergon spp., а также пузырчатки Utricularia spp. (stygia, intermedia, minor). За исключением затопленных гелофитов все остальные компоненты являются необязательными. Utricularia часто покрывают более 20%, а плавающие мхи в основном намного меньше 60%, однако в Польше и странах Балтии покрытие мхами местами выше.
Подводная растительность в основном состоит из затопленных частей гелофитов, как живых, так и мертвых, и плавающих сфагновых и других мхов, таких как напр. Scorpidium scorpioides, Warnstorfia fluitans, Limprichtia revolvens, Calliergon spp., а также пузырчатки Utricularia spp. (stygia, intermedia, minor). За исключением затопленных гелофитов все остальные компоненты являются необязательными. Utricularia часто покрывают более 20%, а плавающие мхи в основном намного меньше 60%, однако в Польше и странах Балтии покрытие мхами местами выше.
Полет тихий и слабый. Держится зарослей трав. Проводит большую часть времени сидя или слабо летая в зарослях осоки или другом подобном укрытии. В очень жаркие дни они уходят в тенистую глубину растительности до полудня, выбираясь в солнечные верхние зоны ближе к вечеру. Если температура выше 15°C и нет ветра, могут быть активны в дни с сильным туманом, частично облачной погодой или даже под небольшим дождем.
При низкой плотности особей, взрослых обнаружить сложно, так как они обычно остаются в густой растительности, прячутся за стеблями и не проявляют особого желания летать, перелетают низко от стебля к стеблю независимо от погоды. Заметить их можно чаще всего лишь вспугнув, когда они перелетают на небольшое расстояние. Предположительно низкая подвижность довольно короткокрылых имаго является одной из причин редкости вида.
Весь жизненный цикл, включая развитие, поиск пищи, отдых, ночевку и размножение, проходит в пространственно ограниченной среде обитания. Плотная привязка к определенному упоминается несколькими авторами. Например, сообщается, что 92% особей оставались в пределах личиночного развития, в том числе на расстоянии до 10 м от нее, и почти ни одна особь не была обнаружена более чем в 100 м от зоны откладывания яиц. Помеченные особи за 10 дней переместились максимум на 25 м.
В исключительных случаях имаго встречаются вдали от подходящей среды обитания. Однако регистрации особей в необычных местах, в том числе удаленных от мест размножения, могут быть результатом уноса ветром.
При низкой плотности особей, взрослых обнаружить сложно, так как они обычно остаются в густой растительности, прячутся за стеблями и не проявляют особого желания летать, перелетают низко от стебля к стеблю независимо от погоды. Заметить их можно чаще всего лишь вспугнув, когда они перелетают на небольшое расстояние. Предположительно низкая подвижность довольно короткокрылых имаго является одной из причин редкости вида.
Весь жизненный цикл, включая развитие, поиск пищи, отдых, ночевку и размножение, проходит в пространственно ограниченной среде обитания. Плотная привязка к определенному упоминается несколькими авторами. Например, сообщается, что 92% особей оставались в пределах личиночного развития, в том числе на расстоянии до 10 м от нее, и почти ни одна особь не была обнаружена более чем в 100 м от зоны откладывания яиц. Помеченные особи за 10 дней переместились максимум на 25 м.
В исключительных случаях имаго встречаются вдали от подходящей среды обитания. Однако регистрации особей в необычных местах, в том числе удаленных от мест размножения, могут быть результатом уноса ветром.
В благоприятную погоду имаго пребывают в верхних частях растительности до и в течение ночи. Ближе к вечеру они менее активны, хотя еще может происходить кормление, а вечером остаются практически неактивными. Ближе к вечеру, особенно в безветренную и солнечную погоду, они греются на верхних частях растений, ловя последние лучи солнца. Сообщается, что имаго сползают в нижние части растительности после захода солнца., однако это не правило. В Северной Польше многочисленные имаго оставались на ночь точно на той же высоте, что и во время заката, то есть в самой верхней четверти растительности, всего на 1–7 см ниже кончиков растений.
Питается мелкими насекомыми, в основном двукрылыми, но также и микрочешуекрылыми (Elachista sp.). Кормовая активность, хотя и сосредоточена во второй половине дня, продолжается весь день. Основными хищниками поедающими этот вид, скорее всего, являются пауки (Araneidae, Tetragnathidae) и лягушки (Rana Lessonae), которые многочисленны во многих местах обитания вида. Однако, несмотря на то, что имаго довольно часто попадались в паутине, не наблюдалось ни одного успешного нападения лягушки. Молодых особей иногда поедают лютки и стрелки. Тенеральные особи иногда заражаются личинками водяных клещей (Arrenurus sp.), что значительно снижает летные способности имаго.
Обладает плохой способностью к расселению, селится небольшими, разбросанными колониями. Численность сокращается и в настоящее время вымер во многих западных частях своего ареала. Сопутствующие виды включают Coenagrion puella, C. pulchellum, Aeshna juncea, Libellula quadrimaculata и Sympetrum danae.
Питается мелкими насекомыми, в основном двукрылыми, но также и микрочешуекрылыми (Elachista sp.). Кормовая активность, хотя и сосредоточена во второй половине дня, продолжается весь день. Основными хищниками поедающими этот вид, скорее всего, являются пауки (Araneidae, Tetragnathidae) и лягушки (Rana Lessonae), которые многочисленны во многих местах обитания вида. Однако, несмотря на то, что имаго довольно часто попадались в паутине, не наблюдалось ни одного успешного нападения лягушки. Молодых особей иногда поедают лютки и стрелки. Тенеральные особи иногда заражаются личинками водяных клещей (Arrenurus sp.), что значительно снижает летные способности имаго.
Обладает плохой способностью к расселению, селится небольшими, разбросанными колониями. Численность сокращается и в настоящее время вымер во многих западных частях своего ареала. Сопутствующие виды включают Coenagrion puella, C. pulchellum, Aeshna juncea, Libellula quadrimaculata и Sympetrum danae.
Активность начинается рано утром. Через 2 часа после восхода солнца многие особи могут быть уже активны, а самцы начинать искать самок. Такая ранняя активность наблюдалась и при относительно низкой температуре 15,5°С с легким туманом после туманной и довольно теплой ночи на севере Польши.
Активность имаго меняется погодой, т. е. совместным воздействием температуры, влажности, инсоляции и ветра. Вид активен, в том числе репродуктивно, уже при температуре от 15 до 18°C. Особи избегают сильной инсоляции, особенно палящего солнечного света в средние часы очень теплых дней с температурой выше 25°С, в то время как утренний и вечерний солнечный свет, хотя и не обязательный, кажется для нехалении благоприятным. Облачность, похоже, не помеха для вида. При соответствующей температуре имаго остаются активными даже в морось или небольшой дождь. Во время сильного дождя они могут оставаться в верхних частях растительности, малоподвижными, с опущенными вниз брюшками. Однако очень ограничивающим фактором для вида является ветер. Особи держатся низко в растительности или ищут защищенное от ветра место уже при его умеренной силе. В одной и той же местности заметны разительные различия в активности между открытыми и защищенными от ветра местами.
Наиболее благоприятными погодными условиями для имаго являются безветрие или очень слабый ветер, умеренная температура, частичная или легкая облачность при приглушенном солнечном свете, т.е. отсутствие сильной инсоляции и предшествующие теплые ночи. При ярком солнечном свете в жаркие дни или в солнечные дни с прохладным ветром нехаления в основном прячется среди растительности.
Активность имаго меняется погодой, т. е. совместным воздействием температуры, влажности, инсоляции и ветра. Вид активен, в том числе репродуктивно, уже при температуре от 15 до 18°C. Особи избегают сильной инсоляции, особенно палящего солнечного света в средние часы очень теплых дней с температурой выше 25°С, в то время как утренний и вечерний солнечный свет, хотя и не обязательный, кажется для нехалении благоприятным. Облачность, похоже, не помеха для вида. При соответствующей температуре имаго остаются активными даже в морось или небольшой дождь. Во время сильного дождя они могут оставаться в верхних частях растительности, малоподвижными, с опущенными вниз брюшками. Однако очень ограничивающим фактором для вида является ветер. Особи держатся низко в растительности или ищут защищенное от ветра место уже при его умеренной силе. В одной и той же местности заметны разительные различия в активности между открытыми и защищенными от ветра местами.
Наиболее благоприятными погодными условиями для имаго являются безветрие или очень слабый ветер, умеренная температура, частичная или легкая облачность при приглушенном солнечном свете, т.е. отсутствие сильной инсоляции и предшествующие теплые ночи. При ярком солнечном свете в жаркие дни или в солнечные дни с прохладным ветром нехаления в основном прячется среди растительности.
Как правило, является довольно медлительной стрекозой, активность связана почти исключительно с одной и той же средой обитания, которую совместно используют тенеральные, полувзрослые и взрослые особи обоих полов. Оперативно соотношение полов несколько смещено в сторону самцов и меняется в течение дня с увеличением числа самок. Присады и полеты на короткие расстояния, «прыжки» с одной присады на другую - наиболее типичные элементы поведения. Угол между осью тела и более или менее вертикальной присадой, частота полетов и агрессивного поведения, а также интенсивность репродуктивного поведения указывают на уровень активности. Установлено, что при температуре 10-10,5°С и в полной темноте особи не реагировали ни на свет фонарика, ни на почти касание руки наблюдателя, несмотря на хорошее освещение. Днем при температуре ниже 13°С, а также при несколько более высокой температуре при прохладном ветре особи вообще не летали. В случае опасности они часто перелазят на другую сторону тонкого стебля растения, чтобы скрыть тело от наблюдателя. Такое поведение иногда также проявляется при более высоких температурах и регулярно у тенеральных особей независимо от температуры.
При низких температурах или во время сильных дождей стрекозы сидят в позе с телом почти параллельным опоре или под углом до 30°. Преобладание углов от 30 до 60° в отдельной популяции свидетельствует о довольно низком общем уровне активности, которая может заключаться в небольшом количестве демонстраций, редких коротких перелетах и начале репродуктивного поведения. Диапазон 45-90° со средними значениями между 60 и 80° типичен для активных особей. Преобладание углов 70-90° в популяции наблюдается ближе к вечеру после погожего дня, несмотря на низкую летную активность, особи, вероятно, в этой позе ловят поздние солнечные лучи.
При низких температурах или во время сильных дождей стрекозы сидят в позе с телом почти параллельным опоре или под углом до 30°. Преобладание углов от 30 до 60° в отдельной популяции свидетельствует о довольно низком общем уровне активности, которая может заключаться в небольшом количестве демонстраций, редких коротких перелетах и начале репродуктивного поведения. Диапазон 45-90° со средними значениями между 60 и 80° типичен для активных особей. Преобладание углов 70-90° в популяции наблюдается ближе к вечеру после погожего дня, несмотря на низкую летную активность, особи, вероятно, в этой позе ловят поздние солнечные лучи.
Полеты обычно короткие и низкие, только в исключительных случаях нехалении поднимаются над растительностью, даже в периоды максимальной активности. В начальной и конечной фазах активности дистанции полета редко превышают 10-15 см, тогда как на пике они обычно колеблются от 10 см до 1 м, однако полеты самцов в поисках самок и некоторых копулирующих пар могут быть более продолжительными. Полеты редко длятся дольше 10 сек., за исключением поисковых полетов самцов, которые могут продолжаться до 2 мин. Очень короткие полеты вверх и вниз могут служить вертикальным перемещением среди растительности. Однако в начальной и конечной фазах активности такое перемещение также может быть достигнуто путем переползания вверх и вниз.
Агрессивное поведение очень редко заканчивается физическим контактом. Чаще всего оно ограничивается проявлениями угрозы сидящего на присаде или летящего самца в ответ на приближение другого самца или тандема, как своего вида, так и чужого (например, Coenagrion sp.). Демонстрация угрозы включает поднятие VIII-X сегментов брюшка с заметным голубым пятном, самцы на присаде также усиливают ее частично расправленными и иногда трепещущими крыльями. В наиболее энергичной форме демонстрация угрозы находит свое выражение в зависании лицом к лицу, иногда включающем «танцы», т. е. внезапные подъемы и опускания малой амплитуды.
Демонстрация отказа самок заключается в изгибании брюшка снизу, у сидящих особей также в расправлении крыльев, в полете иногда «танцы», подобные танцам между самцами, но с большей амплитудой до 20 см. У молодых самок, окрашенных как самцы, демонстрация угрозы может заменять демонстрацию отказа.
Агрессивное поведение очень редко заканчивается физическим контактом. Чаще всего оно ограничивается проявлениями угрозы сидящего на присаде или летящего самца в ответ на приближение другого самца или тандема, как своего вида, так и чужого (например, Coenagrion sp.). Демонстрация угрозы включает поднятие VIII-X сегментов брюшка с заметным голубым пятном, самцы на присаде также усиливают ее частично расправленными и иногда трепещущими крыльями. В наиболее энергичной форме демонстрация угрозы находит свое выражение в зависании лицом к лицу, иногда включающем «танцы», т. е. внезапные подъемы и опускания малой амплитуды.
Демонстрация отказа самок заключается в изгибании брюшка снизу, у сидящих особей также в расправлении крыльев, в полете иногда «танцы», подобные танцам между самцами, но с большей амплитудой до 20 см. У молодых самок, окрашенных как самцы, демонстрация угрозы может заменять демонстрацию отказа.
Самцы пробираются сквозь сплетение стеблей в поисках самок. Когда представители обоих полов встречаются, они выполняют своеобразный танец, летая вверх и вниз друг перед другом. После захвата самки происходит передача спермы.
Через 2,5 часа после восхода солнца как в Польше, так и в Баден-Вюртемберге, были обнаружены первые тандемы. Копуляции начинаются через 2,5-3,25 ч после восхода солнца. Поскольку копуляции, особенно начавшиеся утром, длятся чаще всего долго, брачные кольца накапливаются в последующие часы, достигая численного пика поздним утром. На двух участках в северной Польше последние образования тандемов произошли через 2 и 2,25 часа после полудня, а последние два тандема наблюдались в 15:20.
Для формирования сопряженного колеса может потребоваться 50 минут, а само спаривание длится от 1 до 3 часов. Самка обычно откладывает яйца в одиночку в гниющие стебли и листья осоки. Яйцекладка происходит в юго-западной части европейского ареала с 11 до 17 часов.
Через 2,5 часа после восхода солнца как в Польше, так и в Баден-Вюртемберге, были обнаружены первые тандемы. Копуляции начинаются через 2,5-3,25 ч после восхода солнца. Поскольку копуляции, особенно начавшиеся утром, длятся чаще всего долго, брачные кольца накапливаются в последующие часы, достигая численного пика поздним утром. На двух участках в северной Польше последние образования тандемов произошли через 2 и 2,25 часа после полудня, а последние два тандема наблюдались в 15:20.
Для формирования сопряженного колеса может потребоваться 50 минут, а само спаривание длится от 1 до 3 часов. Самка обычно откладывает яйца в одиночку в гниющие стебли и листья осоки. Яйцекладка происходит в юго-западной части европейского ареала с 11 до 17 часов.
Репродуктивное поведение напоминает поведение других стрелок и включает следующие элементы: a) ожидание самок в местах откладки яиц или активный поиск партнера самцами в густой растительности, б) захват самки и формирование тандема, в) приглашение к совокуплению с последующим перемещением спермы у самца, которое длится несколько секунд, и «генитальное прикосновение» самки, все эти элементы, выполняются в тандеме на присаде, г) совокупление с тремя стадиями, включая быстрое «пинание» самца, происходящее последовательно с довольно регулярным ритмом, функция которого остается неизвестной; копуляции могут продолжаться от 0,5 до 4,5 ч, продолжительность уменьшается в течение дня, д) яйцекладка, в основном менее чем через 1-3 мин после окончания копуляции, обычно без охраны самцом и продолжающаяся до 2 ч; яйца откладываются в гниющие стебли и листья осоки, плавающие на поверхности воды.
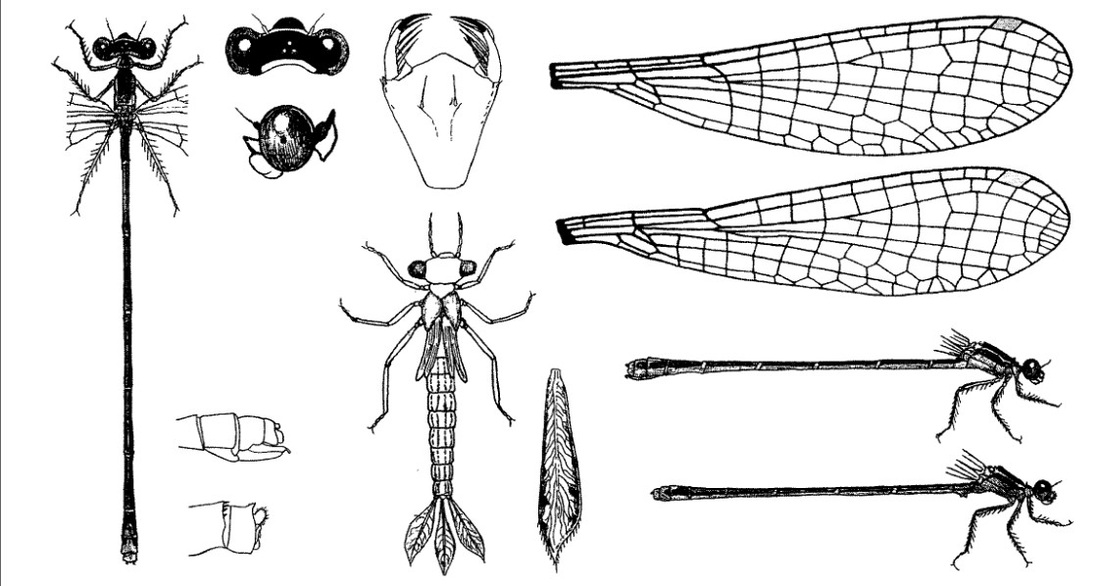
Длина личинки и экзувия не превышает 13 мм, жабры закруглены на конце, но расширены до небольшой точки посередине. Яйца и личинки могут пережить короткие периоды частичного высыхания во влажных сфагновых подушках. Исключительный белорусский случай, возможно, можно отнести к этой категории.
Личинки встречаются в мелких болотистых водоемах с обязательным присутствием осоки. Личинки живут между отмершими стеблями осоки, пузырчатки и мхами. В этой микросреде мало конкуренции и хищников.
Для воды места обитания личинок характерны: а) глубина от нескольких до 30 (40) см; за единственным исключением - влажный сфагнум без открытой воды; б) окраска часто слегка коричневато-желтоватая, обусловленная гуминовыми кислотами; в) быстрое прогревание и высокие температуры, которые в солнечные летние дни могут достигать 30°С. Даже в прохладные и ветреные летние дни в некоторых местах северной Польши температура воды превышала температуру воздуха на 1,5-7°С, причем разница была наиболее заметна между сентябрем и началом ноября, т. е. в период интенсивного роста личинок. Зимой температура во впадинах, покрытых снегом, держится около 0°С, и вода замерзает только в суровых условиях; (г) довольно низкий уровень рН, в пределах от 3,3 до 7,2, в Польше в основном 4,0-5,5; е) низкая жесткость, от менее 0,1 до 5,7, но чаще всего не более 0,5°dH; ж) малотрофные, чаще всего описываемые как олиготрофные или мезотрофные, однако на многих участках воды могут быть обозначены как дистрофные.
Встречаемость N. speciosa в солоноватых водах (менее 0,5-0,7% солености) является исключением, она зарегистрирована в одном месте в Финляндии.
В целом, типичная среда обитания представляет собой стоячую, неглубокую, теплую и довольно кислую воду. Она бедна карбонатами и другими электролитами и может подвергаться локальному влиянию минеральных грунтовых вод.
Личинки встречаются в мелких болотистых водоемах с обязательным присутствием осоки. Личинки живут между отмершими стеблями осоки, пузырчатки и мхами. В этой микросреде мало конкуренции и хищников.
Для воды места обитания личинок характерны: а) глубина от нескольких до 30 (40) см; за единственным исключением - влажный сфагнум без открытой воды; б) окраска часто слегка коричневато-желтоватая, обусловленная гуминовыми кислотами; в) быстрое прогревание и высокие температуры, которые в солнечные летние дни могут достигать 30°С. Даже в прохладные и ветреные летние дни в некоторых местах северной Польши температура воды превышала температуру воздуха на 1,5-7°С, причем разница была наиболее заметна между сентябрем и началом ноября, т. е. в период интенсивного роста личинок. Зимой температура во впадинах, покрытых снегом, держится около 0°С, и вода замерзает только в суровых условиях; (г) довольно низкий уровень рН, в пределах от 3,3 до 7,2, в Польше в основном 4,0-5,5; е) низкая жесткость, от менее 0,1 до 5,7, но чаще всего не более 0,5°dH; ж) малотрофные, чаще всего описываемые как олиготрофные или мезотрофные, однако на многих участках воды могут быть обозначены как дистрофные.
Встречаемость N. speciosa в солоноватых водах (менее 0,5-0,7% солености) является исключением, она зарегистрирована в одном месте в Финляндии.
В целом, типичная среда обитания представляет собой стоячую, неглубокую, теплую и довольно кислую воду. Она бедна карбонатами и другими электролитами и может подвергаться локальному влиянию минеральных грунтовых вод.
Обычно развитие унивольтинное и зимовка на личиночной стадии. Однако 10-20% популяции могут быть семивольтинными. Доля двулетних личинок, вероятно, зависит от весенней и осенней температуры воды.
Период отрождения имаго приходится на середину мая и начало июля, пик - на середину июня, причем половина личинок превращается в имаго через 10-36 дней после начала отрождений. Превращение личинки в имаго происходит в течение дня с пиком утром или днем, в зависимости от погоды. Экзувии встречаются на растительности на высоте 10-20 см над водой. Однако в Польше и Прибалтике почти все экзувии были обнаружены значительно ниже, от 2 до 10 см над водой и лишь в исключительных случаях выше. Стебли осоки в довольно плотно заросших местах являются основным субстратом для отрождения.
Период отрождения имаго приходится на середину мая и начало июля, пик - на середину июня, причем половина личинок превращается в имаго через 10-36 дней после начала отрождений. Превращение личинки в имаго происходит в течение дня с пиком утром или днем, в зависимости от погоды. Экзувии встречаются на растительности на высоте 10-20 см над водой. Однако в Польше и Прибалтике почти все экзувии были обнаружены значительно ниже, от 2 до 10 см над водой и лишь в исключительных случаях выше. Стебли осоки в довольно плотно заросших местах являются основным субстратом для отрождения.

♂ Фото © Семён Левый, Докшицкий р-н, Витебская обл.
Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
4. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
5. Smallshire D., Swash A. "Sedgling. Nehalennia speciosa" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 120-121.
6. Kalniņš M. Bernard R., Miķelsone I. "Protected Aquatic Insects of Latvia – Nehalennia speciosa (Charpentier, 1840) (Odonata: Coenagrionidae)" / Latvijas entomologs 2011, 50: 41-54.
7. Bernard R., Wildermuth H. "Nehalennia speciosa (Charpentier, 1840) in Europe: a case of a vanishing relict (Zygoptera: Coenagrionidae)" / Odonatologica 34(4): 335-378. December 1, 2005.
8. Wildermuth H. "Merkblätter Arten – Libellen – Nehalennia speciosa" / Schweizerische Arbeitsgemeinschaft für Libellenschutz, CSCF info fauna, Neuenburg und Bundesamt für Umwelt, Bern, 2013. 5 S.
9. Mauersberger R. "Zwerglibelle – Nehalennia speciosa (Charpentier 1840)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 60-61.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
4. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
5. Smallshire D., Swash A. "Sedgling. Nehalennia speciosa" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 120-121.
6. Kalniņš M. Bernard R., Miķelsone I. "Protected Aquatic Insects of Latvia – Nehalennia speciosa (Charpentier, 1840) (Odonata: Coenagrionidae)" / Latvijas entomologs 2011, 50: 41-54.
7. Bernard R., Wildermuth H. "Nehalennia speciosa (Charpentier, 1840) in Europe: a case of a vanishing relict (Zygoptera: Coenagrionidae)" / Odonatologica 34(4): 335-378. December 1, 2005.
8. Wildermuth H. "Merkblätter Arten – Libellen – Nehalennia speciosa" / Schweizerische Arbeitsgemeinschaft für Libellenschutz, CSCF info fauna, Neuenburg und Bundesamt für Umwelt, Bern, 2013. 5 S.
9. Mauersberger R. "Zwerglibelle – Nehalennia speciosa (Charpentier 1840)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 60-61.
