БАБКА ЖЕЛТОПЯТНИСТАЯ
Somatochlora flavomaculata Vander Linden, 1825

♂ Фото © Владимир Бондарь, Могилевский р-н
Somatochlora flavomaculata Vander Linden, 1825 (бабка желтопятнистая)
Среднего размера, чуть меньше S. metallica. Глаза ярко-зеленые. На лбу 2 желтых пятна, в углу около глаз. Грудь металлического изумрудно-зеленого цвета с двумя желтыми продолговатыми отметинами внизу. Брюшко черноватое с желтыми метками по бокам сегментов, наибольшими на II-III и узким желтым кольцом вокруг основания сегмента III (у самца неполное). Сегменты брюшка IV-VIII с боковыми темно-желтыми (иногда оранжевыми) пятнами, I-III (либо II-III) с ярко-желтыми. Желтые отметки на брюшке у самцов меньше, чем у самок, с возрастом становятся темнее у обоих полов и особенно трудно различимы у старых самцов. Крылья с желтым оттенком у основания.
♂: Брюшко перетянуто на сегменте III и слегка вздуто на VI-VII (у Cordulia aenea на VII-VIII); кончик держится в полете менее приподнятым. Верхние анальные придатки параллельны друг другу от основания до конца, самые кончики загнуты кверху (у C. aenea довольно короткие и тупые).
♀: Боковые желтые пятна брюшка крупнее и соединены друг с другом, образуя непрерывную и широкую зигзагообразную полосу. Брюшко имеет параллельные стороны и заметный «шип» (генитальную пластинку), который короче сегмента IX (длиннее у S. metallica), и заметно выступающий ниже сегмента IX. Сама генитальная пластинка оканчивается двумя короткими широко закругленными лопастями. Костальная жилка желтая.
Неполовозрелые особи имеют коричневые глаза и ярко-желтые пятна на теле.
Лет: конец мая - конец августа.
Длина 45-54 мм, брюшко 34-40 мм, заднее крыло 32-38 мм.
♂: Брюшко перетянуто на сегменте III и слегка вздуто на VI-VII (у Cordulia aenea на VII-VIII); кончик держится в полете менее приподнятым. Верхние анальные придатки параллельны друг другу от основания до конца, самые кончики загнуты кверху (у C. aenea довольно короткие и тупые).
♀: Боковые желтые пятна брюшка крупнее и соединены друг с другом, образуя непрерывную и широкую зигзагообразную полосу. Брюшко имеет параллельные стороны и заметный «шип» (генитальную пластинку), который короче сегмента IX (длиннее у S. metallica), и заметно выступающий ниже сегмента IX. Сама генитальная пластинка оканчивается двумя короткими широко закругленными лопастями. Костальная жилка желтая.
Неполовозрелые особи имеют коричневые глаза и ярко-желтые пятна на теле.
Лет: конец мая - конец августа.
Длина 45-54 мм, брюшко 34-40 мм, заднее крыло 32-38 мм.

♂ Фото © Владимир Бондарь, Могилевский р-н
Населяет широкий спектр стоячих и медленно текущих водоемов, преимущественно теплых и густозаросших или заболоченных.
Территориальные самцы патрулируют места размножения, часто на уровне глаз наблюдателя. Однако кормятся взрослые особи в периферийных районах, для спаривания укрываясь на деревьях.
Среда размножения - неглубокие торфянистые воды в топях, тростниковых канавах, болотах с буйной растительностью и рядом с лесом; иногда медленнотекущие стоки и каналы. Площади открытой воды, как правило, небольшие и уменьшаются из-за разрастания надводной растительности.
У представителей семейства существуют несколько тактик поиска партнера, то есть активный поиск партнера путем сканирования больших территорий в медленном полете с глубоким проникновением в густую растительность и поиском откладывающих яйца самок или фиксированное патрулирование. Самцы этого вида использует обе тактики, применение каждой из которых зависит от структуры среды обитания. На однородно структурированных участках откладки яиц площадью более 0,05 га самцы обычно медленно пересекают открытые участки и активно ищут прилетающих или откладывающих яйца самок. Однако в местообитаниях с неоднородной ландшафтной структурой и разрозненными местами яйцекладки самцы становятся территориальными. Они занимают фиксированные участки, предпочтительно в непосредственной близости от кустов, деревьев и других вертикальных элементов, выступающих из травянистой растительности.
Территориальные самцы патрулируют места размножения, часто на уровне глаз наблюдателя. Однако кормятся взрослые особи в периферийных районах, для спаривания укрываясь на деревьях.
Среда размножения - неглубокие торфянистые воды в топях, тростниковых канавах, болотах с буйной растительностью и рядом с лесом; иногда медленнотекущие стоки и каналы. Площади открытой воды, как правило, небольшие и уменьшаются из-за разрастания надводной растительности.
У представителей семейства существуют несколько тактик поиска партнера, то есть активный поиск партнера путем сканирования больших территорий в медленном полете с глубоким проникновением в густую растительность и поиском откладывающих яйца самок или фиксированное патрулирование. Самцы этого вида использует обе тактики, применение каждой из которых зависит от структуры среды обитания. На однородно структурированных участках откладки яиц площадью более 0,05 га самцы обычно медленно пересекают открытые участки и активно ищут прилетающих или откладывающих яйца самок. Однако в местообитаниях с неоднородной ландшафтной структурой и разрозненными местами яйцекладки самцы становятся территориальными. Они занимают фиксированные участки, предпочтительно в непосредственной близости от кустов, деревьев и других вертикальных элементов, выступающих из травянистой растительности.

♂ Фото © Владимир Бондарь, Могилевский р-н
Обычно репродуктивно активные самцы обустраивают свои территории в местах, где они могут встретить восприимчивых самок. В большинстве случаев первичное рандеву соответствует месту откладки яиц, тогда как регулярное образование пар вдали от воды является скорее исключением. Одно из таких исключений встречается у S. flavomaculata, самцы которых часто занимают территории вдали от воды, но не исключительно, так как надводные территории также регистрируются регулярно, хотя и реже.
В неоднородной среде самцы S. flavomaculata регулярно занимают фиксированные места вдали от воды, примыкающие к вертикальным элементам ландшафта, и в меньшей степени также у воды, т. е. в местах откладки яиц. Территории как над сушей, так и над водой обычно патрулируются непрерывными фиксированными полетами. Эти места служат местами встреч с самками, где начинается совокупление.
В неоднородной среде самцы S. flavomaculata регулярно занимают фиксированные места вдали от воды, примыкающие к вертикальным элементам ландшафта, и в меньшей степени также у воды, т. е. в местах откладки яиц. Территории как над сушей, так и над водой обычно патрулируются непрерывными фиксированными полетами. Эти места служат местами встреч с самками, где начинается совокупление.

♂ Фото © Виктор Рындевич, Минский р-н
Результаты семилетнего исследования в неоднородной болотистой среде Центральной Европы с разбросанными местами откладки яиц показали, что места встреч менялись в течение лётного сезона как по местоположению, так и по качеству. В начале периода размножения территории формировались почти исключительно на суше. В дальнейшем произошел значительный сдвиг территорий над сушей к территориям над водой, а к концу лётного сезона практически все территории оказались над водой. Участки с заросшими лужами также были привлекательными для обустройства территорий для размножения даже в начале лётного сезона. Когда лужи высыхали в первой половине периода размножения, эти места больше не использовались в качестве мест встреч. Однако смещения в сторону надводных территорий в этой ситуации не наблюдалось. Мелкомасштабный перенос территорий также был связан с температурой окружающей среды. Территориальные самцы в основном летали при температуре окружающей среды от 20 до 34°С. При температуре до 28°C все самцы патрулировали при ярком солнечном свете, но когда температура повышалась, они постепенно перемещали места патрулирования в тень, по-видимому, из соображений терморегуляции. В промежутках они патрулировали частично на солнце и частично в тени. Похоже, что стратегия поиска партнера у S. flavomaculata характеризуется значительной фенотипической пластичностью по отношению ко времени и пространству.

♀ Фото © Денис Китель, Пружанский р-н, Брестская обл.
Интересное исследование проводилось с 2002 по 2008 год на болоте площадью 15 га в кантоне Цюрих, Швейцария. Формирование тандемов на территориях и последующее совокупление в воздухе происходило очень быстро и наблюдалось всего 5 раз над сушей и дважды над водой. Большинство из 194 совокупляющихся пар не были обнаружены до тех пор, пока не были замечены летящими в положении "колеса". Обычно пары кружили довольно низко над растительностью или над прудами и лужами в течение нескольких минут, прежде чем сесть на травянистую растительность или перелететь на куст или дерево. Точное время копуляции не было определено, так как пары часто перемещались и терялись из виду, но за несколькими парами, сидящими в низкой растительности, наблюдали в течение 15-20 мин, прежде чем они перелетели дальше. Окончание копуляции наблюдалось дважды. После того, как партнеры разделились, они оставались сидеть на растительности довольно близко друг к другу, а затем улетели один за другим. Яйцекладка происходила без сопровождения самца и в основном скрыто в густой растительности. За период исследования было замечено 10 самок, откладывающих яйца.

♂ Фото © Семён Левый, Житковичский р-н, Гомельская обл.
Известно, что закрепленные места патрулирования у самцов S. flavomaculata размещаются как над сушей (наземные), так и над водой (водные), и эти места служат местами встреч с самками. Неоднократно наблюдалось формирование пар на обоих типах территорий, свидетельствуя о том, что самцы регулярно поджидают самок как в местах откладки яиц, так и вне их. В исключительных случаях совокупляющиеся пары S. flavomaculata встречаются даже в лесных промежутках, вдали от водоемов.
Места спаривания меняются с наземных на водные в течение периода размножения не случайно, а регулярно, по крайней мере, на неоднородном участке с множеством мелких и разрозненных мест размножения. Причины сезонного перехода от преимущественно наземных к преимущественно водным территориям малопонятны. Возможно, самки определяют подходящие места для откладывания яиц во время посещения водных или болотных участков и приближаются к этим местам все ближе и ближе по ходу сезона. Кроме того, дополнительные совокупления могут больше быть не нужны для самок в конце сезона. Следовательно, они стараются избегать преследований самцов и украдкой прилетают к местам откладки яиц. Таким образом, к концу лётного сезона наземные территории постепенно теряют свою ценность в качестве мест встреч, и самцы реагируют на изменение, перемещая свои территории к воде.
Места спаривания меняются с наземных на водные в течение периода размножения не случайно, а регулярно, по крайней мере, на неоднородном участке с множеством мелких и разрозненных мест размножения. Причины сезонного перехода от преимущественно наземных к преимущественно водным территориям малопонятны. Возможно, самки определяют подходящие места для откладывания яиц во время посещения водных или болотных участков и приближаются к этим местам все ближе и ближе по ходу сезона. Кроме того, дополнительные совокупления могут больше быть не нужны для самок в конце сезона. Следовательно, они стараются избегать преследований самцов и украдкой прилетают к местам откладки яиц. Таким образом, к концу лётного сезона наземные территории постепенно теряют свою ценность в качестве мест встреч, и самцы реагируют на изменение, перемещая свои территории к воде.

♂ Фото © Максим Мясников, Дзержинский р-н, Минская обл.
Нимфа. Личинки очень похожи на личинок S. metallica, отличаются от них отсутствием шипов на затылке, более длинным и более узким латеральным шипом на IX сегменте, очень маленьким или совершенно отсутствующим латеральным шипом на VIII сегменте и меньшими размерами. Голова короткая; темя вздутое, округлое, с каждой стороны его темные отметины; глаза маленькие, слегка выпуклые, полусферические, расположены в передних углах головы; затылок широкий, без выдающихся бугорков, с 8 шероховатыми продольными полосами, окруженными яйцевидными плоскими пространствами, задний край почти прямой. Подбородок с 5-6 зубцами на дистальном крае боковых лопастей. Переднегрудь с приподнятым задним краем, короткая, значительно уже головы. Ноги длиннее и тоньше, чем у личинок S. metallica, бедра задней пары доходят до VII сегмента; голени, а также и бедра этой пары, темные, с 1 светлым у основания и с 4 светлыми вблизи вершины кольцами; первый членик лапок задней пары на 1/3 короче второго. Брюшко овальное, широкое, с усеченной вершиной; X сегмент маленький, узкий; дорсальные шипы на III-IX; на VI-IX сегментах они острые, с искривленными назад вершинами, длинные, заходят за нижележащие сегменты; латеральный шип на VIII сегменте или очень маленький, меньше 1/6 длины края этого сегмента, или совсем отсутствует; шип на IX сегменте острый, плоский, длинный, длиннее 1/2 внешнего края этого сегмента, заходит за Х сегмент. Анальная пирамида такой же длины, как IX и Х сегменты, взятые вместе; дорсальный придаток немного короче церков, треугольный, сильно расширенный у основания, островершинный; церки короткие, заостренные; церкоиды такой же длины, как дорсальный придаток, с внезапно заостряющейся вершиной.

♂ Фото © Владимир Бондарь, Могилевский р-н
Личинки живут главным образом в маленьких ручейках, проточных канавах, в заводях рек, но встречаются также в озерах, прудах, торфяных болотах, карьерах торфяных разработок и т. п. Регулярно заселяются особо плотные заросли телореза в небольших водоемах, канавах или отделенных от озер бухтах. Обычно на местообитание приходится всего несколько экзувий.
Фаза развития продолжительностью не менее 2 лет. Отрождения имаго начинаются с конца мая и продолжаются до конца июня.
Фаза развития продолжительностью не менее 2 лет. Отрождения имаго начинаются с конца мая и продолжаются до конца июня.

♂ Фото © Владимир Бондарь, Могилевский р-н

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
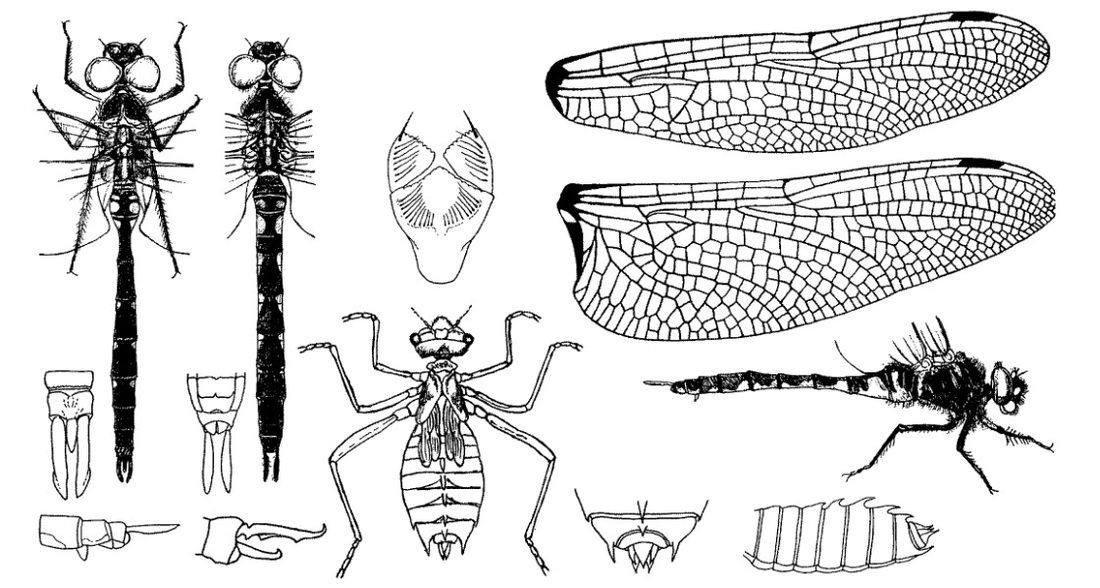
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Smallshire D., Swash A. "Yellow-spotted Emerald. Somatochlora flavomaculata" / Europe's Dragonflies: A field guide to the damselflies and dragonflies" Princeton University Press, 2020. pp. 240-241.
8. Wildermuth H. "Season and temperature dependent location of mating territories in Somatochlora flavomaculata in a heterogeneous environment (Odonata: Corduliidae)" / International Journal of Odonatology 12 (2) 2009: 181-193, pl. III DOI: 10.1080/13887890.2009.9748338
9. Mauersberger R., Brauner O. "Gefleckte Smaragdlibelle – Somatochlora flavomaculata (Vander Linden 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 104-105.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Smallshire D., Swash A. "Yellow-spotted Emerald. Somatochlora flavomaculata" / Europe's Dragonflies: A field guide to the damselflies and dragonflies" Princeton University Press, 2020. pp. 240-241.
8. Wildermuth H. "Season and temperature dependent location of mating territories in Somatochlora flavomaculata in a heterogeneous environment (Odonata: Corduliidae)" / International Journal of Odonatology 12 (2) 2009: 181-193, pl. III DOI: 10.1080/13887890.2009.9748338
9. Mauersberger R., Brauner O. "Gefleckte Smaragdlibelle – Somatochlora flavomaculata (Vander Linden 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 104-105.
