СТРЕКОЗА ОБЫКНОВЕННАЯ
Sympetrum vulgatum Linnaeus, 1758

© Виктор Рындевич, Минский р-н
Sympetrum vulgatum Linnaeus, 1758 (стрекоза обыкновенная)
Верх глаз коричневый. Черная поперечная полоса, расположенная между теменем и лбом, спускается книзу вдоль внутреннего края глаз. Задний край переднегруди с большим, почти вертикальным выступом, несущим бахромку из длинных волосков. Бока груди тусклые с более широкими черными полосками на швах. Брюшко имеет черные отметки вдоль вершин сегментов VIII и IX. Крылья бесцветные, только в основании задних небольшое желтоватое пятнышко. Птеростигмы от желтых до красновато-коричневых. Ноги черные, с желтой продольной полосой снаружи.
♂: Грудь практически без меток, коричневая с красноватым оттенком. Брюшко красное, с черной боковой полосой и с крошечными черными пятнами на сегментах III-VII, каждое из которых окружено желтым кольцом. Брюшко слегка перетянутое, сегменты V-VIII немного расширены. Наружные ветви генитальных крючков заострены; верхние анальные придатки почти сплошь светлые.
♀: Бока груди желто-бурые, с прямоугольником из тонких черных линий в центре. Брюшко желто-охристое с прерывистыми мелкими черными полосками по бокам. С возрастом желтый цвет в окраске тускнеет; некоторые самки становятся красноватыми, как самцы. Генитальная пластинка видна, торчит вертикально вниз и заострена.
Неполовозрелые похожи на ярких самок.
♂: Грудь практически без меток, коричневая с красноватым оттенком. Брюшко красное, с черной боковой полосой и с крошечными черными пятнами на сегментах III-VII, каждое из которых окружено желтым кольцом. Брюшко слегка перетянутое, сегменты V-VIII немного расширены. Наружные ветви генитальных крючков заострены; верхние анальные придатки почти сплошь светлые.
♀: Бока груди желто-бурые, с прямоугольником из тонких черных линий в центре. Брюшко желто-охристое с прерывистыми мелкими черными полосками по бокам. С возрастом желтый цвет в окраске тускнеет; некоторые самки становятся красноватыми, как самцы. Генитальная пластинка видна, торчит вертикально вниз и заострена.
Неполовозрелые похожи на ярких самок.

♀ Фото © Семён Левый
Похож на S. striolatum, но немного меньше его, у половозрелых самцов брюшко более красноватое и более суженное в средних сегментах, хотя и не в такой степени, как у самцов S. sanguineum. Грудь без двух желтых пятен на боках, в отличие от S. striolatum. Черная поперечная линия на вершине лба проходит вниз по внутренней стороне глаз, в отличие от S. striolatum, у которого нет ее латерального расширения или, самое большее, имеется нечеткий мазок.
Длина 35-40 мм, брюшко 24-28 мм, заднее крыло 24-29 мм.
Лет: начало июля - начало октября.
Длина 35-40 мм, брюшко 24-28 мм, заднее крыло 24-29 мм.
Лет: начало июля - начало октября.

♀ Фото © Семён Левый
Населяет широчайший спектр стоячих и застойных водоемов с обильной водной растительностью. Населяет очень широкий спектр стоячих и медленно текущих водоемов, таких как пруды, озера, заводи, затопленные карьеры и канавы. Кроме того, встречается в значительно меньшей степени и плотности на низких, переходных и верховых болотах. Видимо, предпочитает обширные мелководья и зоны заиления с тростниковыми зарослями и зарослями осоки на берегах. Избегает только рек с быстрым течением или стоячих вод, полностью окруженных деревьями и кустарниками, сильно затененных или очень кислых. Часто синтопичен с S. striolatum, но по сравнению с ним, как правило, обитает в водоемах с несколько более поздними стадиями сукцессии и при пышной подводной и надводной растительности, а иногда и с несколько более низкой температурой воды.
На тропинках в пойменных лугах наиболее часто можно встретить самцов. Другие виды рода в этих местах бывают редко. В период созревания самцов и самок иногда можно увидеть вдали от воды, например, по залежам и опушкам леса.
Может разогревать свои летательные мышцы за счет активного дрожания крыльев и максимального пребывания тела на солнце, чтобы он оставался активным в полете при температуре окружающей среды не ниже 12°C.
На тропинках в пойменных лугах наиболее часто можно встретить самцов. Другие виды рода в этих местах бывают редко. В период созревания самцов и самок иногда можно увидеть вдали от воды, например, по залежам и опушкам леса.
Может разогревать свои летательные мышцы за счет активного дрожания крыльев и максимального пребывания тела на солнце, чтобы он оставался активным в полете при температуре окружающей среды не ниже 12°C.

♀ Фото © Семён Левый, Кобринский р-н, Брестская обл.
Способность вида к распространению считается очень высокой, так как он может быстро заселять вновь образованные водоемы. Иногда мигрирует, но менее склонный к крупномасштабному расселению, чем другие виды.
Хотя для Беларуси не свойственны массовые миграции не только этого вида, но и стрекоз вообще, представляется интересным описание явления, несколько уникального и наблюдаемого на юге Западной Сибири в Барабинской лесостепи. Уникальность такой миграции заключалась в том, что она совершалась в тандемах.
В 2010 г. миграции тандемов S. vulgatum и S. danae на юге Западной Сибири были особенно интенсивными. Для возникновения таких перелетов требуется два обстоятельства. Первое – высокая численность этих видов в текущем году. Второе - сочетание, по крайней мере, трех погодных факторов: высокой, не ниже 20°С, температуры, наличия солнца и ветра не менее 3, но не более 10-12 м/с. Оптимальная скорость ветра 5–8 м/с, направление ветра не имеет значения. Наиболее важным и интригующим обстоятельством является то, что стрекозы при этом всегда летят против ветра: в 80% наблюдавшихся случаев строго навстречу ветру и в 20% под небольшим углом к нему с отклонением в ту или иную сторону. Соответствующие погодные условия в конце лета и осенью создаются далеко не каждый день, поэтому в массовых перелетах тандемов нет периодичности.
Хотя для Беларуси не свойственны массовые миграции не только этого вида, но и стрекоз вообще, представляется интересным описание явления, несколько уникального и наблюдаемого на юге Западной Сибири в Барабинской лесостепи. Уникальность такой миграции заключалась в том, что она совершалась в тандемах.
В 2010 г. миграции тандемов S. vulgatum и S. danae на юге Западной Сибири были особенно интенсивными. Для возникновения таких перелетов требуется два обстоятельства. Первое – высокая численность этих видов в текущем году. Второе - сочетание, по крайней мере, трех погодных факторов: высокой, не ниже 20°С, температуры, наличия солнца и ветра не менее 3, но не более 10-12 м/с. Оптимальная скорость ветра 5–8 м/с, направление ветра не имеет значения. Наиболее важным и интригующим обстоятельством является то, что стрекозы при этом всегда летят против ветра: в 80% наблюдавшихся случаев строго навстречу ветру и в 20% под небольшим углом к нему с отклонением в ту или иную сторону. Соответствующие погодные условия в конце лета и осенью создаются далеко не каждый день, поэтому в массовых перелетах тандемов нет периодичности.

♀ Фото © Семён Левый
В утренние часы самцы и самки держатся раздельно, иногда образуя большие скопления вдоль березовых колков, лесополос, зарослей тростника и рослого травостоя вдоль полевых дорог. При благоприятной погоде они начинают кормиться примерно с 8 ч утра, а после 9 ч начинают образовывать первые тандемы. С 10 до 11 ч происходит массовое образование тандемов, и при достаточном прогреве воздуха, солнечной и ветреной погоде множество образовавшихся пар начинает полет против ветра. В него вовлекаются не все тандемы, некоторая часть из них летит для яйцекладки на любой ближайший водоем. Однако чем ближе к осени, тем большее число пар вместо перемещения к ближайшему водоему устремляется в дальний перелет навстречу ветру. Однонаправленное движение тандемов стрекоз в большинстве наблюдавшихся случаев достигало максимума в середине дня и после 14 ч начинало ослабевать, полностью прекращаясь в 15–16 ч. После этого тандемы распадаются, самцы и самки кормятся, отдыхают и ночуют независимо друг от друга. На следующее утро при благоприятной погоде тандемы образуются заново.

♂ Фото © Семён Левый, Борисовский р-н, Минская обл.
В разное время и в разных местах “плотность” потока летящих тандемов стрекоз менялась в больших пределах. В 2010 г. согласованное по направлению движение тандемов симпетрумов впервые было зарегистрировано 17 августа. В середине дня при ясной погоде, юго-западном ветре 5–7 м/с и температуре воздуха 26°С наблюдалось движение спаренных стрекоз навстречу ветру с интенсивностью в 40±8.8 пар (25–52) через полосу длиной 10 м за 10 мин наблюдений, или 4 пары/мин.
Все последующие дни почти до конца сентября, когда погода отвечала перечисленным выше условиям, лёт тандемов симпетрумов наблюдался как на этом участке, так и в радиусе десятков километров от него.
30 августа был зарегистрирован самый масштабный из всех наблюдавшихся перелетов тандемов симпетрумов. Происходил он в те же часы, при ясной погоде, температуре 23-25°С и северо–северо-западном ветре силой в среднем 5 м/с. Наблюдение велось вдоль береговой линии оз. Фадиха длиной 1 км. Максимальный по интенсивности лёт наблюдался между 11 ч 40 мин и 14 ч. Стрекозы в массе летели почти строго на север под небольшим углом к ветру. Одновременно в поле зрения находились до 200 тандемов. Подсчитывать летящих насекомых из-за их большого количества было трудно, и время каждого учета пришлось сократить до 5 мин. В серии из десяти учетов в период максимального лёта за 5 мин 10-метровую линию пересекло 722±94.8 тандема (610–870), или 144.4 пар/мин. Соответственно, за 1 мин через 1 км местности пролетало примерно 14440 тандемов стрекоз, за 1 ч - более 866 тыс., а за 2 ч 20 мин наиболее активной фазы этой миграции пролетело примерно 2 млн. пар.
В течение сентября интенсивность однонаправленных перелетов спаренных симпетрумов в целом ослабевала, но они наблюдались до конца месяца. Хотя размножение S. vulgatum и S. danae продолжалось до середины октября, полеты тандемов наблюдались все реже, и образующиеся пары независимо от направления ветра и поведения других тандемов чаще всего летели для яйцекладки в сторону ближайшего водоема.
Все последующие дни почти до конца сентября, когда погода отвечала перечисленным выше условиям, лёт тандемов симпетрумов наблюдался как на этом участке, так и в радиусе десятков километров от него.
30 августа был зарегистрирован самый масштабный из всех наблюдавшихся перелетов тандемов симпетрумов. Происходил он в те же часы, при ясной погоде, температуре 23-25°С и северо–северо-западном ветре силой в среднем 5 м/с. Наблюдение велось вдоль береговой линии оз. Фадиха длиной 1 км. Максимальный по интенсивности лёт наблюдался между 11 ч 40 мин и 14 ч. Стрекозы в массе летели почти строго на север под небольшим углом к ветру. Одновременно в поле зрения находились до 200 тандемов. Подсчитывать летящих насекомых из-за их большого количества было трудно, и время каждого учета пришлось сократить до 5 мин. В серии из десяти учетов в период максимального лёта за 5 мин 10-метровую линию пересекло 722±94.8 тандема (610–870), или 144.4 пар/мин. Соответственно, за 1 мин через 1 км местности пролетало примерно 14440 тандемов стрекоз, за 1 ч - более 866 тыс., а за 2 ч 20 мин наиболее активной фазы этой миграции пролетело примерно 2 млн. пар.
В течение сентября интенсивность однонаправленных перелетов спаренных симпетрумов в целом ослабевала, но они наблюдались до конца месяца. Хотя размножение S. vulgatum и S. danae продолжалось до середины октября, полеты тандемов наблюдались все реже, и образующиеся пары независимо от направления ветра и поведения других тандемов чаще всего летели для яйцекладки в сторону ближайшего водоема.

♂ Фото © Денис Китель.
Во всех наблюдавшихся случаях массового лёта тандемов среди них в незначительном числе попадались и одиночные особи обоих полов, летящих в том же направлении, что и поток сцепленных пар. Очень редко среди тандемов встречались “триплеты” - сцепленные в одну цепочку три стрекозы: летящий первым самец удерживал анальными придатками брюшка второго самца за его голову, который, в свою очередь, удерживал самку. Маневренность в полете тандемов и “триплетов” очень высокая, и отлов их сачком не менее затруднителен, чем одиночных стрекоз в том же потоке. Большинство тандемов летело на высоте 1-3 м, хотя некоторые двигались над самой землей или, наоборот, летели на высоте 10 м и более. При движении над водоемами, потенциально привлекательными в качестве репродуктивных стаций, отдельные тандемы делали кратковременные остановки для яйцекладки, после которой всегда продолжали полет в прежнем направлении. Доля таких пар была очень невысока. Например, в прибрежной зоне оз. Фадиха, представляющей собой благоприятную репродуктивную стацию для исследуемых стрекоз, из 10 тыс. тандемов, пролетевших через 10-метровый сектор наблюдения 30 августа, только 12 пар сделали остановку для откладки яиц.
Вскрытие 20 самок S. vulgatum из пойманных тандемов показало, что их брюшки забиты яйцами. В среднем количество зрелых яиц у одной самки 1130±235 штук и еще примерно 100-200 более мелких незрелых яиц находилось в яйцеводах, т.е. репродуктивная система стрекоз находилось в оптимальном для яйцекладки состоянии.
Вскрытие 20 самок S. vulgatum из пойманных тандемов показало, что их брюшки забиты яйцами. В среднем количество зрелых яиц у одной самки 1130±235 штук и еще примерно 100-200 более мелких незрелых яиц находилось в яйцеводах, т.е. репродуктивная система стрекоз находилось в оптимальном для яйцекладки состоянии.

♀ Фото © Виталий Некрашевич, г. Брест
Массовое однонаправленное перемещение тандемов стрекоз представляет собой своеобразный и высоко адаптивный вариант расселения, при котором обеспечивается максимально полное распределение на местности откладываемых яиц. Учитывая, что симпетрумы откладывают яйца не только в водоемы, но и в любые понижения рельефа, заливаемые следующей весной талыми водами, представляется целесообразной стратегия тотального прочесывания местности летящими тандемами с “осеменением” яйцами всех потенциальных биотопов. Яйца откладываются каждой парой небольшими порциями по мере их готовности к кладке, а так как у каждой пары время такой готовности наступает индивидуально, то в целом на всю массу летящих тандемов процесс должен получаться достаточно равномерным: одна пара отложила порцию яиц здесь, другая - там и так далее. В движении тандемы не сбиваются в более-менее плотные стаи, как это бывает при других типах миграций, а движутся в одном общем направлении широким фронтом, захватывающим целые ландшафты. А в качестве фактора, задающего направление всему потоку мигрантов, выступает ветер, навстречу которому летят стрекозы. Поскольку направление ветра может меняться, меняется и курс полетов тандемов, т.е. стрекозы неоднократно пересекают одну и ту же территорию в разных направлениях, что делает распределение яиц на местности более плотным и равномерным. Массовые передвижения тандемов по описанному выше сценарию, строго говоря, вряд ли можно считать миграциями. Правильнее их называть репродуктивными кочевками. В любом случае на территориях с нестабильной обводненностью такая стратегия способна поддерживать максимально выровненную плотность популяций и более стабильную численность вида.

♀ Фото © Татьяна Алешка, Лепельский р-н, Витебская обл.
Предпочитает низкие и часто светлые присады, с которых можно наблюдать за потенциальной добычей или партнерами. Взрослые самцы большую часть дня проводят у береговой линии. Стычки за территории обычно происходят утром. Однако в принципе, у самцов этого вида сравнительно мало меж- и внутривидовых конфликтов. Самки обычно проводят день, охотясь на солнечных лугах или залежах, и прилетают непосредственно к водоему только для размножения.
Спаривание происходит в основном около полудня, длится всего несколько минут, начинается в полете и обычно заканчивается сидя на растительности. Пары иногда отмечаются далеко от воды. Попытки спаривания с другими видами рода происходят регулярно, чему, вероятно, способствует относительно неспециализированный копулятивный аппарат и схожие репродуктивные среды обитания внутри рода. Экзофитная яйцекладка производится парами сразу после совокупления. Самец при кладке яиц сопровождает самку, часто в конце кладки покидает ее, но не улетает, а держится рядом, летая вокруг или позади самки. Регулярно можно увидеть группы из более чем 10 пар, откладывающих яйца одновременно.
Спаривание происходит в основном около полудня, длится всего несколько минут, начинается в полете и обычно заканчивается сидя на растительности. Пары иногда отмечаются далеко от воды. Попытки спаривания с другими видами рода происходят регулярно, чему, вероятно, способствует относительно неспециализированный копулятивный аппарат и схожие репродуктивные среды обитания внутри рода. Экзофитная яйцекладка производится парами сразу после совокупления. Самец при кладке яиц сопровождает самку, часто в конце кладки покидает ее, но не улетает, а держится рядом, летая вокруг или позади самки. Регулярно можно увидеть группы из более чем 10 пар, откладывающих яйца одновременно.

Фото © Семён Левый, г. Брест.
Яйца откладываются в прибрежный ил, сырую землю у воды и и на подводную растительность, достигающую поверхности воды, причем самка каждый раз ударяет брюшком о субстрат и из полового отверстия выходят комочками влажные и липкие яйца, соединяющиеся в воде в кучку. Яйца часто распределяются по большой площади. Размер кладки яиц сильно различается и составляет в среднем около 800 яиц. Яйца продолговато-овальные, желтоватого цвета, матовые, имеют среднюю длину 0,50 мм и среднюю ширину 0,37 мм.
Зимует на стадии яйца. Яйца могут переживать обезвоживание. Личинки вылупляются из яиц следующей весной при температуре не ниже 15-18°.
Зимует на стадии яйца. Яйца могут переживать обезвоживание. Личинки вылупляются из яиц следующей весной при температуре не ниже 15-18°.

♀ Фото © Семён Левый, Борисовский р-н, Минская обл.
Нимфа. Крупные личинки от темно-серых до красновато-коричневых, часто серо-зеленые, с пестро разрисованным слабо волосатым телом. Голова широкая; на лбу узкая черная полоска и близ основания антенн светло-желтый рисунок, на котором обыкновенно выделяются два Т-образных черных пятна; 3-6-й членики антенн темные в основной и светлые в дистальной части; затылок с темными пятнами и тонкой светлой продольной срединной полоской, имеющейся и по середине темени; задние углы затылка округлые, с 3 грубыми большими щетинками, задний край его обрамлен короткими щетинками. Переднегрудь с тонкой светлой продольной срединной полоской. Ноги всех трех пар с 2 темными полосками. Брюшко дорсально с тонкой светлой продольной срединной полоской, обычно хорошо видной только на сегментах между чехлами крыльев; по обе стороны светлой полоски 2 черноватые или томно-зеленые полосы, контуры которых не резки; иногда все брюшко светлое, у таких особей все сегменты испещрены темными пятнами, составляющими продольные ряды. Дорсальный придаток анальной пирамиды широкий у основания, островершинный, приблизительно на 1/3 короче церков; церки короче длины IX и Х сегментов, взятых вместе, узкие, островершинные; церкоиды в 2 раза короче церков, слабо изогнутые внутрь; все отростки волосатые.

♂ Фото © Семён Левый, Житковичский р-н, Гомельская обл.
Личинки живут в стоячих и слабо текущих водах: заводях рек, озерах, разливах родников, болотцах, лужах, карьерах, но предпочитают заводи рек и озер с более чистой водой, более глубоким дном и сильно заросшие макрофитами (Elodea canadensis, Stratiotes aloides, Nymphaea candida, Nuphar luteum, Ceratophyllum и др.). Они живут на водных растениях или закапываются в ил, развиваются быстро. Питаются в основном зоопланктоном. Могут сосуществовать при умеренной плотности ихтиофауны, и этот вид регулярно встречается в рыбных прудах, однако большое количество свидетельств размножения и развития поступает из водоемов, свободных от рыбы.
В целом, эмбриональное и личиночное развитие менее гибко, чем у S. striolatum. Цикл развития одногодичный, моновольтинный. Впрочем, из-за эвритопности и обширностью арела этот вид имеет гибкий унивольтинный или семивольтинный жизненный цикл, зависящий от условий развития.
Метаморфоз (окрыление имаго) часто происходит прямо над береговой линией на подходящей растительности. Период дополнительного питания молодых имаго, во время которого они широко (иногда на несколько километров) разлетаются от водоемов, занимает 2-3 недели, после чего насекомые приступают к размножению. Вид слабо привязан к местам своего выплода.
В целом, эмбриональное и личиночное развитие менее гибко, чем у S. striolatum. Цикл развития одногодичный, моновольтинный. Впрочем, из-за эвритопности и обширностью арела этот вид имеет гибкий унивольтинный или семивольтинный жизненный цикл, зависящий от условий развития.
Метаморфоз (окрыление имаго) часто происходит прямо над береговой линией на подходящей растительности. Период дополнительного питания молодых имаго, во время которого они широко (иногда на несколько километров) разлетаются от водоемов, занимает 2-3 недели, после чего насекомые приступают к размножению. Вид слабо привязан к местам своего выплода.

♀ Фото © Виталий Некрашевич, г. Брест

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
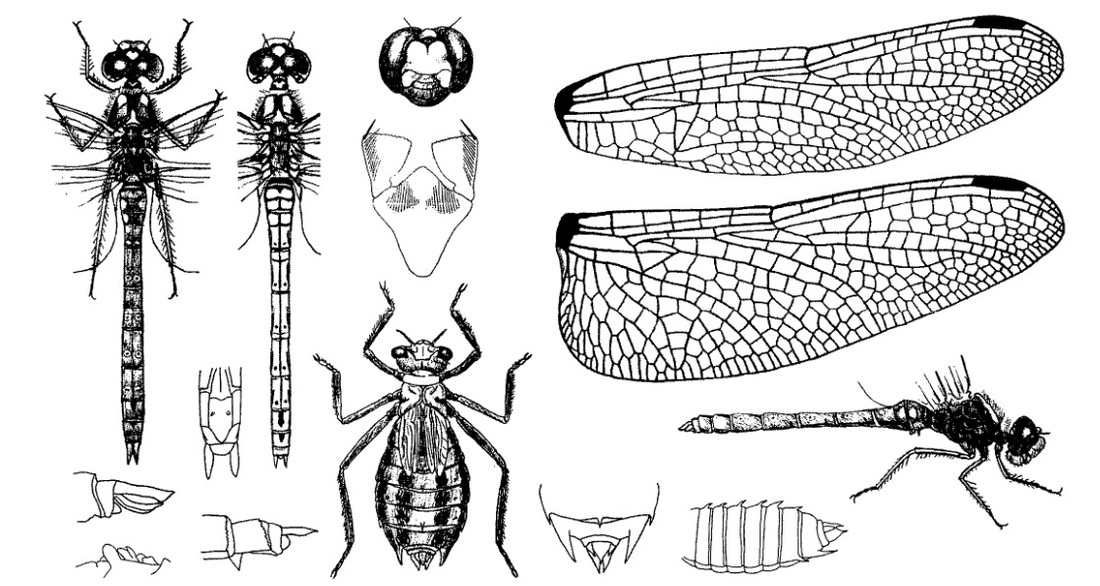
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Moustached Darter. Sympetrum vulgatum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 304-305.
9. Ott J., Koch K. "Sympetrum vulgatum (Linnaeus, 1758) Gemeine Heidelibelle" / im: Brockhaus et all. Atlas der Libellen Deutschlands (Odonata). Libellula Supplement 14: 334-337. 2015.
10. Попова О. Н., Харитонов А. Ю. "Массовые репродуктивные кочевки стрекоз рода Sympetrum (Odonata, Libellulidae)" / Зоологический журнал, 2013, том 92, № 8, с. 893–900.
11. Petzold F., Kruse M., Mauersberger R. "Gemeine Heidelibelle – Sympetrum vulgatum (Linnaeus 1758)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 148-149.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Moustached Darter. Sympetrum vulgatum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 304-305.
9. Ott J., Koch K. "Sympetrum vulgatum (Linnaeus, 1758) Gemeine Heidelibelle" / im: Brockhaus et all. Atlas der Libellen Deutschlands (Odonata). Libellula Supplement 14: 334-337. 2015.
10. Попова О. Н., Харитонов А. Ю. "Массовые репродуктивные кочевки стрекоз рода Sympetrum (Odonata, Libellulidae)" / Зоологический журнал, 2013, том 92, № 8, с. 893–900.
11. Petzold F., Kruse M., Mauersberger R. "Gemeine Heidelibelle – Sympetrum vulgatum (Linnaeus 1758)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 148-149.
