КОРОМЫСЛО ГОЛУБОЕ или ЧЕШУЙЧАТОЕ
Aeshna caerulea Ström, 1783

♂ Photo © Volkmar Wagner / Wikimedia Commons. CC BY-SA 3.0
Aeshna caerulea Ström, 1783 (коромысло голубое или чешуйчатое)
Согласно новым данным, имеются большие сомнения относительно нахождения вида в Беларуси, особенно в Гомельской и Могилевской областях. Ранее вид указывался в обзоре стрекоз Беларуси (Buczyński et al., 2006) на основании литературных данных (Писаненко, 1985 - для Витебской, Гомельской и Могилевской обл.; Кипенварлиц, 1933-34 - для Витебской обл.). Вместе с тем, вид указывается для Латвии и Эстонии, так что вероятность подтверждения вида для республики остается самой высокой на верховых болотах Витебской обл. Тем более, что для Европы не считая реликтовые послеледниковые популяции в горных районах, указывается, что вид встречается в основном выше 55° широты (т. е. севернее линии Лынтупы-Бешенковичи-Лиозно). В любом случае, остро необходимы современные данные о наличии вида в республике.
Голова относительно небольшая; глаза соприкасаются друг с другом вдоль короткой линии. Тело пестрое, голова зеленоватая, на лбу полный Т-образный черный рисунок. Шов между лбом и наличником черный. Грудь бурая или коричневая; на боках две косые полосы; впереди предплечевые полосы очень узкие и не полные. На груди сверху имеются голубые пятнышки у основания крыльев. Жилкование крыльев черное. Костальная жилка коричневатая, а не ярко-желтая, как у A. juncea. Птеростигма черноватая. При посадке принимает вертикальную позу со свисающим вниз брюшком. При холодной погоде голубой цвет сменяется серым.
♂: Глаза голубые сверху, коричневые снизу. Светлые предплечевые полосы очень короткие. Две косые полосы на боках груди голубоватые. Брюшко в основном голубое, с крупными парными пятнами; отсутствие желтых меток является диагностическим. Анальный треугольник из двух ячеек. Ребрышко на тыльной стороне верхних анальных придатков слабо зазубренное.
♀: Глаза коричневатые. Основная окраска коричневая, но встречаются и голубые формы, рисунок на брюшке такой же, как у самца, но все основные светлые метки желтые и более мелкие. Светлые предплечевые полосы очень короткие, но чаще отсутствуют. Две косые полосы на боках груди желтоватые. Яйцеклад короткий, задний конец его не заходит даже за конец последнего сегмента брюшка.
Стрекоза среднего размера, по длине тела похожая на A. mixta. У взрослого самца брюшко черное с обширным рисунком, состоящим преимущественно из парных голубых отметин на дорсальной поверхности брюшка. В отличие от других представителей семейства с голубыми пятнами, у них нет зеленых или желтых отметин на брюшке; таким образом, A. caerulea можно отличить от более крупного и гораздо более обычного A. juncea, который может встречаться в тех же местах (однако необходимо тщательное изучение, поскольку это различие не очевидно в полете). Доплечевые полосы на коричневато-черной груди тонкие или отсутствуют, по бокам груди имеется пара узких светлых полос. Самки более коричневатые, также имеют рисунок парных отметин вдоль брюшка, но они варьируются от тускло-бежевого до бледно-голубого, как у неполовозрелых самцов, и в исключительных случаях бледно-беловатые или ярко-голубые. Локально голубые и коричневые формы самок могут встречаются одинаково часто.
Лет: конец мая - начало сентября.
Длина 54-64 мм, брюшко 42-48 мм, заднее крыло 37-41 мм.
♂: Глаза голубые сверху, коричневые снизу. Светлые предплечевые полосы очень короткие. Две косые полосы на боках груди голубоватые. Брюшко в основном голубое, с крупными парными пятнами; отсутствие желтых меток является диагностическим. Анальный треугольник из двух ячеек. Ребрышко на тыльной стороне верхних анальных придатков слабо зазубренное.
♀: Глаза коричневатые. Основная окраска коричневая, но встречаются и голубые формы, рисунок на брюшке такой же, как у самца, но все основные светлые метки желтые и более мелкие. Светлые предплечевые полосы очень короткие, но чаще отсутствуют. Две косые полосы на боках груди желтоватые. Яйцеклад короткий, задний конец его не заходит даже за конец последнего сегмента брюшка.
Стрекоза среднего размера, по длине тела похожая на A. mixta. У взрослого самца брюшко черное с обширным рисунком, состоящим преимущественно из парных голубых отметин на дорсальной поверхности брюшка. В отличие от других представителей семейства с голубыми пятнами, у них нет зеленых или желтых отметин на брюшке; таким образом, A. caerulea можно отличить от более крупного и гораздо более обычного A. juncea, который может встречаться в тех же местах (однако необходимо тщательное изучение, поскольку это различие не очевидно в полете). Доплечевые полосы на коричневато-черной груди тонкие или отсутствуют, по бокам груди имеется пара узких светлых полос. Самки более коричневатые, также имеют рисунок парных отметин вдоль брюшка, но они варьируются от тускло-бежевого до бледно-голубого, как у неполовозрелых самцов, и в исключительных случаях бледно-беловатые или ярко-голубые. Локально голубые и коричневые формы самок могут встречаются одинаково часто.
Лет: конец мая - начало сентября.
Длина 54-64 мм, брюшко 42-48 мм, заднее крыло 37-41 мм.
Населяет болота, озера, заводи рек. Согласно данным из Британии и Ирландии, размножается на больших открытых участках в неглубоких болотных лужах, по крайней мере с небольшим количеством мха Sphagnum. В Шотландии вид предпочитает небольшие неглубокие водоемы площадью 1-8 м² и глубиной 10-15 см. Надводная растительность скудная, в основном Eriophorum sp. Субстрат представлял собой мягкий супообразный детрит разной толщины. Важным фактором также было присутствие Sphagnum cuspidatum на участках с открытой водой.
Ключевые участки представляют собой исходные местообитания со значительными популяциями, сохраняющимися в течение многих лет. Взрослые особи, расселившиеся из этих районов, колонизируют вторичные места обитания, где популяции могут сохраняться недолго и зависят от иммиграции особей из исходных мест обитания для пополнения популяции. Подмножество вторичных местообитаний называется латентными местообитаниями, где популяции существуют недолго, но служат полезными ступеньками к новым районам обитания и размножения или другим исходным местам обитания с существующими популяциями. Все эти классы среды обитания для размножения действуют как метапопуляция, взаимодействуя как внутри участков, так и между участками в различных географических масштабах.
Самцы часто сидят на солнце на беловатых валунах, белых стволах берез или бледных моховых кочках Racomithum lanughosum, а при холодной погоде оба пола греются на светлых поверхностях в защищенных местах. Непосредственная близость лесных массивов не является существенной особенностью мест размножения этого вида. Хотя по другим данным, взрослые особи обычно укрываются и кормятся в близлежащих лесных массивах, особенно на полянах в березовых рощах и вдоль ручьев. Однако нет никаких сомнений в том, что лесной покров обеспечивает хорошую среду обитания для укрытия и кормления и, безусловно, улучшает место обитания этого вида.
Ключевые участки представляют собой исходные местообитания со значительными популяциями, сохраняющимися в течение многих лет. Взрослые особи, расселившиеся из этих районов, колонизируют вторичные места обитания, где популяции могут сохраняться недолго и зависят от иммиграции особей из исходных мест обитания для пополнения популяции. Подмножество вторичных местообитаний называется латентными местообитаниями, где популяции существуют недолго, но служат полезными ступеньками к новым районам обитания и размножения или другим исходным местам обитания с существующими популяциями. Все эти классы среды обитания для размножения действуют как метапопуляция, взаимодействуя как внутри участков, так и между участками в различных географических масштабах.
Самцы часто сидят на солнце на беловатых валунах, белых стволах берез или бледных моховых кочках Racomithum lanughosum, а при холодной погоде оба пола греются на светлых поверхностях в защищенных местах. Непосредственная близость лесных массивов не является существенной особенностью мест размножения этого вида. Хотя по другим данным, взрослые особи обычно укрываются и кормятся в близлежащих лесных массивах, особенно на полянах в березовых рощах и вдоль ручьев. Однако нет никаких сомнений в том, что лесной покров обеспечивает хорошую среду обитания для укрытия и кормления и, безусловно, улучшает место обитания этого вида.
Из большинства исследований следует, что взрослые особи проводят большую часть своего времени вдали от мест размножения. Неясно, где именно взрослые особи проводят время кроме периода размножения, но они могут преодолевать приличное расстояние, чтобы найти убежище и подходящую среду для размножения.
Активны в солнечную погоду, самцы низко летают над болотистыми лужами и окружающими пустошами в поисках самок. В отличие от A. juncea, они иногда делают кратковременные посадки при поиске водоемов. Есть сомнения в том, является ли этот вид территориальным. Спаривание происходит вдали от воды, и пары в «положении колеса» можно обнаружить отдыхающими на камнях или низкой растительности. Данных о времени совокупления нет, единственное упоминание гласит, что оно "занимает много минут". Также неизвестно, спариваются ли самки за день до откладывания яиц, как и другие виды семейства.
Размножается в неглубоких торфяных болотных лужах, часто очень маленьких (несколько м²), и на осоковых болотах с обилием болотных мхов. Яйца самки откладывают в одиночку в мягкий торф или мох сфагнум в неглубоких болотных лужах площадью до 30 м² и глубиной воды 20 см и 20 см торфяного ила, хотя обычно выбираются лужи меньшего размера. Яйцекладка длится до 3 минут в любом месте и происходит только в солнечные периоды. Яйца впадают в диапаузу и не развиваются до следующего календарного года с вылуплением в конце апреля или мае, в зависимости от погоды ранней весной.
Активны в солнечную погоду, самцы низко летают над болотистыми лужами и окружающими пустошами в поисках самок. В отличие от A. juncea, они иногда делают кратковременные посадки при поиске водоемов. Есть сомнения в том, является ли этот вид территориальным. Спаривание происходит вдали от воды, и пары в «положении колеса» можно обнаружить отдыхающими на камнях или низкой растительности. Данных о времени совокупления нет, единственное упоминание гласит, что оно "занимает много минут". Также неизвестно, спариваются ли самки за день до откладывания яиц, как и другие виды семейства.
Размножается в неглубоких торфяных болотных лужах, часто очень маленьких (несколько м²), и на осоковых болотах с обилием болотных мхов. Яйца самки откладывают в одиночку в мягкий торф или мох сфагнум в неглубоких болотных лужах площадью до 30 м² и глубиной воды 20 см и 20 см торфяного ила, хотя обычно выбираются лужи меньшего размера. Яйцекладка длится до 3 минут в любом месте и происходит только в солнечные периоды. Яйца впадают в диапаузу и не развиваются до следующего календарного года с вылуплением в конце апреля или мае, в зависимости от погоды ранней весной.
Очень интересна экология вида. Поскольку его ареал фактически приурочен к высоким широтам Северной Европы и высокогорным районам Центральной Европы, он рассматривается как типичный ледниковый реликт, нуждающийся в прохладном субарктическом климате. Однако температурные потребности личинок очень сложны. Молодые личинки теплолюбивы и нуждаются в более или менее постоянной температуре около 20°C, в то время как личинки более старших возрастов обычно переносят более низкие температуры, но предпочитают более высокие температуры для линьки и кормления. Из-за коричневого цвета воды, темного (торфянистого) дна и/или мелководья типичные места размножения вида являются более теплыми местообитаниями даже в холодном климате, что обеспечивает быстрый рост личинок. В отличие от водной микросреды обитания личинок, среда имаго прохладная, суровая и некомфортная для стрекоз. Однако необычное поведение при «загорании на солнце» и физиологическая специализация, особенно физиологическая смена окраски у самцов, делают вид хорошо приспособленным к таким условиям. Таким образом, предпочтение взрослыми особями более низких температур ограничивает распространение вида более прохладным климатом Крайнего Севера, субальпийских и альпийских зон. На более низких границах своего ареала в Центральной Европе, где температуры обычно считаются слишком высокими для вида, он проявляет сильную привязанность к сфагновым болотам, микроклимат которых обычно сходен с холодным континентальным. Поэтому в Беларуси вид может быть обнаружен только на сфагновых болотах.
Имаго, в отличие от личинок (см. ниже), благодаря этологическим, морфологическим и физиологическим приспособлениям хорошо подготовлены к прохладному сырому климату.
Лёт у этого вида не очень устойчив: взрослые особи часто отдыхают на прогретом солнцем субстрате, таком как темная (торфяная) почва, камни или стволы деревьев. Это типичные пользователи присад. Они греются на горизонтальном или более или менее вертикальном субстрате, плотно прижимаются телом к земле и опускают крылья так, что кончики крыльев касаются субстрата. Такое поведение довольно необычно для семейства. Из-за прямого контакта тело стрекозы может поглощать тепло субстрата. Кроме того, опущенные крылья работают как теплица: в зависимости от цвета субстрата температура под крыльями может быть на 7°C выше, чем над ними. Таким образом, воздух под крыльями действует воздушный мешок, предотвращающий потерю тепла груди. Наиболее эффективно это работает при сильном ветре; если место открыто ветру, правая или левая законцовки крыльев направлены против ветра. Это максимально увеличивает площадь крыла, подверженную воздействию ветра; крылья прижимаются к основанию, а замкнутый нагретый воздух оптимально защищен. Из-за этого особого поведения A. caerulea требуется очень мало энергии для разогрева и поддержания подходящей температуры тела, в отличие от постоянно летающих стрекоз. При сильном ветре стрекозы обычно греются на земле, где субстрат наиболее теплый и есть укрытие от ветра.
Лёт у этого вида не очень устойчив: взрослые особи часто отдыхают на прогретом солнцем субстрате, таком как темная (торфяная) почва, камни или стволы деревьев. Это типичные пользователи присад. Они греются на горизонтальном или более или менее вертикальном субстрате, плотно прижимаются телом к земле и опускают крылья так, что кончики крыльев касаются субстрата. Такое поведение довольно необычно для семейства. Из-за прямого контакта тело стрекозы может поглощать тепло субстрата. Кроме того, опущенные крылья работают как теплица: в зависимости от цвета субстрата температура под крыльями может быть на 7°C выше, чем над ними. Таким образом, воздух под крыльями действует воздушный мешок, предотвращающий потерю тепла груди. Наиболее эффективно это работает при сильном ветре; если место открыто ветру, правая или левая законцовки крыльев направлены против ветра. Это максимально увеличивает площадь крыла, подверженную воздействию ветра; крылья прижимаются к основанию, а замкнутый нагретый воздух оптимально защищен. Из-за этого особого поведения A. caerulea требуется очень мало энергии для разогрева и поддержания подходящей температуры тела, в отличие от постоянно летающих стрекоз. При сильном ветре стрекозы обычно греются на земле, где субстрат наиболее теплый и есть укрытие от ветра.
На земле они также находятся в оптимальном положении для ловли добычи: во время сильного ветра воздух почти свободен от мелких насекомых, которые являются предпочтительной добычей в солнечные дни, потому что большинство из них скрываются среди растительности. Лишь немногие летают, но очень близко к наземной растительности. Обычно Aeshnidae не могут охотиться на таких насекомых, поскольку обычно обнаруживают и ловят свою добычу снизу. При сильном ветре жертвы улетают ниже, где их невозможно обнаружить сверху. Но для сидящего A. caerulea, наблюдающего с земли, видны жертвы, летящие низко и даже пролетающие над охотником. Таким образом, даже при сильном ветре успех поимки добычи у вида может быть высоким, когда у других представителей семейства он низкий.
Примечательно, что A. caerulea обычно мало обращает внимания на крупных чешуекрылых. Однако в дни с сильным ветром добычей являются только Macrolepidoptera, которые в таких условиях могут быть единственными летающими насекомыми.
Кроме того, сильный ветер требует много энергии для непрерывно летающих стрекоз, потому что они должны лететь против ветра. Шквалистые ветры делают практически невозможным направленный подлет к летящей над землей добыче; но влияние шквалов ветра уменьшается с уменьшением высоты, поэтому у A. caerulea больше шансов поохотиться на насекомых близко у земли. Обычно в таких ситуациях непрерывно летающие стрекозы прекращают свои усилия через несколько минут, а A. caerulea еще остаются активными. Такое поведение "согревания" также позволяет самцам наблюдать за пролетающими самками.
Примечательно, что A. caerulea обычно мало обращает внимания на крупных чешуекрылых. Однако в дни с сильным ветром добычей являются только Macrolepidoptera, которые в таких условиях могут быть единственными летающими насекомыми.
Кроме того, сильный ветер требует много энергии для непрерывно летающих стрекоз, потому что они должны лететь против ветра. Шквалистые ветры делают практически невозможным направленный подлет к летящей над землей добыче; но влияние шквалов ветра уменьшается с уменьшением высоты, поэтому у A. caerulea больше шансов поохотиться на насекомых близко у земли. Обычно в таких ситуациях непрерывно летающие стрекозы прекращают свои усилия через несколько минут, а A. caerulea еще остаются активными. Такое поведение "согревания" также позволяет самцам наблюдать за пролетающими самками.
При температуре выше 22°C имаго можно наблюдать лишь спорадически. При высокой температуре A. caerulea значительно предпочитает более светлые субстраты для отдыха (например, выбеленное солнцем дерево, сухой Trichophorum cespitosum), которые не нагреваются так сильно, как темные субстраты (например, торф). Но это изменение субстрата, очевидно, мало влияет на предотвращение перегрева, поскольку в жаркие дни A. caerulea ищет более затененные места, в то время как другие Aeshnidae все еще активны на солнце. Избегание перегрева путем поиска затененных мест, вероятно, является примитивной формой терморегуляторного поведения. Кроме такого поведения, у A. caerulea нет других способов избежать перегрева, таких как «положение обелиска» у Gomphidae и Libellulidae. Но предотвращение перегрева путем отлета невыгодно, поскольку снижает успешность спаривания из-за сокращения времени пребывания в местах размножения. Это может иметь определенное значение для самцов, так как они, в отличие от самок, заинтересованы в высокой частоте копуляций. Физиологическое изменение окраски у A. caerulea следует рассматривать как терморегуляторный механизм. При температуре ниже 12°C самцы имеют тусклую окраску (темная фаза), а выше 12°C - ярко-голубую (голубая фаза). Из-за более высокого поглощения излучения самцы в темной фазе прогреваются очень быстро и до более высоких значений, чем самцы голубого окраса; прирост температуры составляет до 7°С под действием солнечного излучения. Это позволяет самцам проявлять активность рано утром и при более низких температурах, чем другие стрекозы. В голубой фазе солнечное излучение отражается и хотя бы в какой-то степени эта окраска защищает самца от перегрева. Самцы могут переносить более высокие температуры и дольше, чем самки, находиться вблизи мест размножения.
Нимфа. Коричневая или темно-коричневая; тело стройное с черными точками, штрихами, светлыми и темными полосами; небольшая. Голова маленькая, несколько плоская на вершине; глаза довольно большие, выдающиеся, грушевидные, сильно вытянутые назад по направлению к средней линии головы; затылок широкий, с 3 блестящими пятнами посредине, с малозаметными блестящими пятнами но боковым сторонам, задний край почти прямой. Переднегрудь сзади выпуклая, на каждой стороне средней линии по 1 блестящему поперечному пятну; задний край приподнятый; боковые выросты небольшие, конические, сморщенные. Ноги средней длины; на бедрах с внешней стороны 2 слабо заметные светлые поперечные полосы. Брюшко длинное, сильно выпуклое, значительно суженное под крыловыми чехлами, темно-коричневое (напоминает брюшко личинок Ae. juncea); со спинной стороны с 2 светлыми варьирующими в ширине проходящими до конца IX сегмента продольными полосами и темной (особенно на VIII и IX сегментах) срединной полосой. Анальная пирамида относительно длинная; островершинные церки немного длиннее IХ и Х сегментов, взятых вместе, и значительно длиннее дорсального придатка; последний относительно широкий, цилиндрический, раздвоенный при основании; церкоиды острые.
Лимнофилы; обитают в илу на дне водоемов с открытой водной поверхностью, в торфяных и топких болотах, болотцах, мелких и небольших прудах, в береговых лужицах. Личинки этого вида не были найдены в проточной воде. О продолжительности фазы личинки и о числе линек мало что известно. Вероятно, им требуется 3 года для развития, но эксперименты по разведению в неволе показывают, что может существовать значительная изменчивость. Отрождение крылатых особей наблюдалось на стеблях пушицы и вереска.
Лимнофилы; обитают в илу на дне водоемов с открытой водной поверхностью, в торфяных и топких болотах, болотцах, мелких и небольших прудах, в береговых лужицах. Личинки этого вида не были найдены в проточной воде. О продолжительности фазы личинки и о числе линек мало что известно. Вероятно, им требуется 3 года для развития, но эксперименты по разведению в неволе показывают, что может существовать значительная изменчивость. Отрождение крылатых особей наблюдалось на стеблях пушицы и вереска.
Эксперименты по выращиванию в различных температурных условиях показывают, что личинки вида в течение первых нескольких недель после вылупления являются теплолюбивыми. Они могут пережить этот период только в том случае, если температура воды примерно постоянна на уровне 21°С или днем и ночью колебалась только между 16 и 21°С. При постоянных температурах 16 и 26°С все личинки 1-го и 2-го возрастов погибли; также при ежедневных колебаниях температуры от 16 до 26°С в первые недели после вылупления личинки не выживали. Эти и некоторые другие эксперименты позволяют предположить, что молодые личинки нуждаются в более или менее постоянной температуре около 20°С.
Большинство личинок выжило при температуре 21°C в период эмбрионального и личиночного развития. Все личинки, содержащиеся при температуре 16°C или 26°C, погибли до 11-го возраста, но когда температура на стадии яйца составляла 21°C, личинки выживали заметно дольше при 16°C, чем при температуре эмбриона 16 или 26°C. Личинки наиболее чувствительны к микроклимату (и другим условиям окружающей среды) в 1-6 возрастах, особенно в возрасте 1-2. В этот период смертность личинок была самой высокой. Личинки старше 6-го возраста сравнительно нечувствительны и наиболее устойчивы ко многим воздействиям окружающей среды.
В опыте личинки меняли свое местоположение в пределах температурного градиента неравномерно. Но все личинки каждой возрастной группы проводили большую часть времени в определенном температурном диапазоне. В самой молодой группе она составляла около 20°С, что являлось оптимальным температурным значением для их развития. Но незадолго до и после линьки и во время пищеварения они предпочитали температуру выше 20°С. Личинки старшего возраста предпочитали другие температуры. Эти эксперименты позволяют предположить, что личинки A. caerulea имеют множество различных температурных требований, которые меняются с возрастом, физиологическим состоянием личинки и сезоном. Полевые наблюдения подтверждают эти результаты.
Большинство личинок выжило при температуре 21°C в период эмбрионального и личиночного развития. Все личинки, содержащиеся при температуре 16°C или 26°C, погибли до 11-го возраста, но когда температура на стадии яйца составляла 21°C, личинки выживали заметно дольше при 16°C, чем при температуре эмбриона 16 или 26°C. Личинки наиболее чувствительны к микроклимату (и другим условиям окружающей среды) в 1-6 возрастах, особенно в возрасте 1-2. В этот период смертность личинок была самой высокой. Личинки старше 6-го возраста сравнительно нечувствительны и наиболее устойчивы ко многим воздействиям окружающей среды.
В опыте личинки меняли свое местоположение в пределах температурного градиента неравномерно. Но все личинки каждой возрастной группы проводили большую часть времени в определенном температурном диапазоне. В самой молодой группе она составляла около 20°С, что являлось оптимальным температурным значением для их развития. Но незадолго до и после линьки и во время пищеварения они предпочитали температуру выше 20°С. Личинки старшего возраста предпочитали другие температуры. Эти эксперименты позволяют предположить, что личинки A. caerulea имеют множество различных температурных требований, которые меняются с возрастом, физиологическим состоянием личинки и сезоном. Полевые наблюдения подтверждают эти результаты.
Высокая тепловая потребность личинок A. caerulea, по-видимому, не соответствует климатическим условиям Крайнего Севера и высокогорных районов. Температура в местах размножения этого вида повышается быстрее и до более высоких значений за счет солнечной радиации, поскольку их глубина составляет всего 10-20 см, они часто заросшие, цвет их воды более или менее коричневый из-за растворенных гуминовых кислот и субстрат часто состоит из темного торфа. Кроме того, днем личинки предпочитают густые мхи (Sphagnum, Drepanocladus), которые достигают поверхности воды и темный цвет которых дополнительно подогревает воду. В холодные ночи только верхние слои воды становятся прохладными, а более глубокие слои остаются теплыми. Кроме того, колебания температуры в верхних слоях воды велики, но по сравнению с температурой окружающей среды вне воды температура более глубоких слоев воды остается на высоком и почти постоянном уровне в пределах одного и того же водоема даже в ночное время, а также в короткие периоды периоды непогоды. В совокупности это приводит к очень подходящему микроклиматическому режиму в местах обитания личинок.
Слишком мелкие водоемы пересыхают в периоды засухи и это может иметь большое локальное влияние на выживаемость личинок. Такие водоемы часто очень маленькие (1х1 м), но есть и большие, более 5 м². Высыхание этих неглубоких водоемов приводит к высокой смертности, в результате чего лишь немногие личинки последнего года жизни вырастают до имаго. Водоемы с более глубокими участками, глубиной более 20 см, предлагают убежище в засуху, однако, является ли это критерием выбора самкой места откладки яиц или просто удачей, пока нет ответа.
Старшие личиночные стадии очень трудно обнаружить, по-видимому, потому, что они ищут другую среду обитания, чем более молодые личинки (которых довольно легко найти) и больше «прячутся» в лужицах по бокам и на дне, а не греются на подводных Sphagnum и Eriophorum spp. Они также являются наименее многочисленной личиночной стадией из-за смертности личинок на предыдущих стадиях.
Слишком мелкие водоемы пересыхают в периоды засухи и это может иметь большое локальное влияние на выживаемость личинок. Такие водоемы часто очень маленькие (1х1 м), но есть и большие, более 5 м². Высыхание этих неглубоких водоемов приводит к высокой смертности, в результате чего лишь немногие личинки последнего года жизни вырастают до имаго. Водоемы с более глубокими участками, глубиной более 20 см, предлагают убежище в засуху, однако, является ли это критерием выбора самкой места откладки яиц или просто удачей, пока нет ответа.
Старшие личиночные стадии очень трудно обнаружить, по-видимому, потому, что они ищут другую среду обитания, чем более молодые личинки (которых довольно легко найти) и больше «прячутся» в лужицах по бокам и на дне, а не греются на подводных Sphagnum и Eriophorum spp. Они также являются наименее многочисленной личиночной стадией из-за смертности личинок на предыдущих стадиях.
Считается, что задержка вылупления защищает самые ранние личиночные стадии, которые наиболее восприимчивы к низким температурам. В течение первых нескольких недель после вылупления личинки теплолюбивы и нуждаются в достаточно постоянной температуре около 20°С; более взрослые личинки более устойчивы. Температурный режим может различаться в разных частях пруда, было замечено, что личинки проникают в более глубокие части водоема в прохладную ночь, но остаются в теплых поверхностных водах в солнечный день.
Личинки цепляются за растительность близко к поверхности воды, оставаясь неподвижными, чтобы замаскироваться. Они быстро и активно реагируют на движение, с готовностью выслеживая или преследуя потенциальную добычу, если она не подошла к ним достаточно близко. Описание их как «ползающих», охотящихся на ощупь, не соответствует тому факту, что у личинок большие глаза, и поэтому они могут быть активными охотниками, использующими зрение, а не осязание. Золотистая окраска личинок является прекрасным камуфляжем на фоне зелено-золотого S. cuspidatum. Как и на взрослых стадиях, личинки поедают любых животных своего размера или меньше, включая сородичей.
Жизненный цикл личинок предполагается от 3 до 4 лет, но может занять еще больше времени в более прохладных местах развития. У личинок предпоследнего возраста вероятно может быть диапауза для обеспечения относительно синхронизированного раннего отрождения имаго в конце мая - конце июня.
Отрождение имаго происходит либо непосредственно на сфагнуме на краю водоема, либо на надводной растительности, например, на осоке Carex spp. В Шотландии отрождение происходит около полудня, начинаясь чуть раньше его и завершаясь всоре после полудня. Эксперименты в неволе показали, что личинки могут завершить свое отрождение за гораздо меньшее время, чем A. juncea, что, возможно, дает им преимущество в продуваемых ветром и открытых местах.
Личинки цепляются за растительность близко к поверхности воды, оставаясь неподвижными, чтобы замаскироваться. Они быстро и активно реагируют на движение, с готовностью выслеживая или преследуя потенциальную добычу, если она не подошла к ним достаточно близко. Описание их как «ползающих», охотящихся на ощупь, не соответствует тому факту, что у личинок большие глаза, и поэтому они могут быть активными охотниками, использующими зрение, а не осязание. Золотистая окраска личинок является прекрасным камуфляжем на фоне зелено-золотого S. cuspidatum. Как и на взрослых стадиях, личинки поедают любых животных своего размера или меньше, включая сородичей.
Жизненный цикл личинок предполагается от 3 до 4 лет, но может занять еще больше времени в более прохладных местах развития. У личинок предпоследнего возраста вероятно может быть диапауза для обеспечения относительно синхронизированного раннего отрождения имаго в конце мая - конце июня.
Отрождение имаго происходит либо непосредственно на сфагнуме на краю водоема, либо на надводной растительности, например, на осоке Carex spp. В Шотландии отрождение происходит около полудня, начинаясь чуть раньше его и завершаясь всоре после полудня. Эксперименты в неволе показали, что личинки могут завершить свое отрождение за гораздо меньшее время, чем A. juncea, что, возможно, дает им преимущество в продуваемых ветром и открытых местах.

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
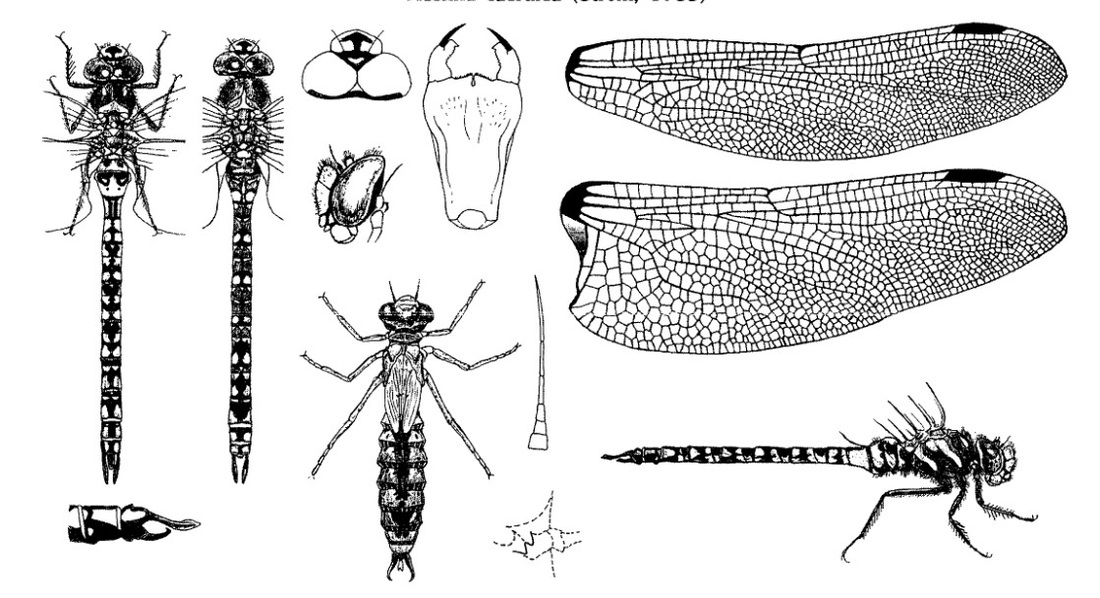
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Китель Д. А. "О сомнительных обнаружениях некоторых видов стрекоз в Беларуси" / Личное сообщение от 02.07.22.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Azure Hawker. Aeshna caerulea" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 146-147.
9. Smallshire D., Swash A. "Azure Hawker Aeshna caerulea" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 102-103.
10. Sternberg K. "Adaptation of Aeshna caerulea (Ström) to the severe climate of its environment (Anisoptera: Aeshnidae)" / Odonatologica 26(4): 439-449. December I, 1997.
11. Willet J. "Species Review 7: The Azure Hawker Aeshna caerulea (Ström)" / Journal of the British Dragonfly Society, Volume 29 No. 1, 2013. pp. 1-19.
12. Batty P. M. "Recent Observations of Aeshna caerulea (Azure Hawker) in Scotland" / Journal of the British Dragonfly, Volume 35 No. 1, 2019. pp. 1-17.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Китель Д. А. "О сомнительных обнаружениях некоторых видов стрекоз в Беларуси" / Личное сообщение от 02.07.22.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Azure Hawker. Aeshna caerulea" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 146-147.
9. Smallshire D., Swash A. "Azure Hawker Aeshna caerulea" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 102-103.
10. Sternberg K. "Adaptation of Aeshna caerulea (Ström) to the severe climate of its environment (Anisoptera: Aeshnidae)" / Odonatologica 26(4): 439-449. December I, 1997.
11. Willet J. "Species Review 7: The Azure Hawker Aeshna caerulea (Ström)" / Journal of the British Dragonfly Society, Volume 29 No. 1, 2013. pp. 1-19.
12. Batty P. M. "Recent Observations of Aeshna caerulea (Azure Hawker) in Scotland" / Journal of the British Dragonfly, Volume 35 No. 1, 2019. pp. 1-17.
