БАБКА ДВУПЯТНИСТАЯ
Epitheca bimaculata Charpentier, 1825

♂ Фото © Денис Китель
Epitheca bimaculata Charpentier, 1825 (бабка двупятнистая)
Относительно крупная стрекоза. Окраска без металлического блеска. Глаза тусклые, сине-зеленые. Лоб оранжевый с черным поперечным пятном. Грудь оливково-бурая, плечевой и первый боковой швы черные. Брюшко длинное, толстое, несколько уплощенное; черное с желто-коричневыми отметинами по бокам, без металлического блеска. Боковые желтые отметки на брюшке более темные и сливающиеся на вершине сегментов I-II, уменьшаются в размерах до незначительных или вовсе отсутствует на сегментах IX-X. Сегменты III-VIII сверху черные или бурые с большими боковыми желтыми пятнами, которые у самки сливаются в продольные полосы. Передний край всех крыльев интенсивно-желтый, впрочем интенсивность желтизны может быть переменной. На задних крыльях при основании по большому черному треугольному пятну (у самки немного крупнее) - отсюда и название "двупятнистая". Ноги очень длинные.
♂ Брюшко приталенное у основания и несколько заостренное к концу. При виде сверху черная дорсальная полоса (с неровными краями) значительно шире, чем у самки. Верхние придатки сдвинуты у основания и изогнуты наружу.
♀ Брюшко значительно шире, чем у самца. Талии нет и примерно с середины брюшко плавно сужается, на конце не такое заостренное, как у самца. Желтые боковые пятна на брюшке более обширные и при виде сверху черная дорсальная полоса значительно уже, чем у самца. Верхние придатки раздвинуты в основании, часто параллельны друг другу.
У неполовозрелых особей глаза коричневые; брюшко ярче, чем у половозрелой взрослой особи, с более обширным охристым и неравномерным черным рисунком на вершине сегментов III-X, как у неполовозрелой Libellula fulva; на крыльях широко распространен золотистый оттенок.
Несколько сходен с двумя более мелкими видами Libellula fulva и L. quadrimaculata. Окраска брюшка у самки или неполовозрелого самца L. fulva похожа на окраску E. bimaculata, но отличается темной линией у основания переднего крыла и темными вершинами крыльев. L. quadrimaculata у обоих полов имеет более или менее развитое черное пятно на узелках всех крыльев.
Длина 55-65 мм, брюшко 39-43 мм, заднее крыло 38-44 мм.
Лет: конец мая - начало июля.
♂ Брюшко приталенное у основания и несколько заостренное к концу. При виде сверху черная дорсальная полоса (с неровными краями) значительно шире, чем у самки. Верхние придатки сдвинуты у основания и изогнуты наружу.
♀ Брюшко значительно шире, чем у самца. Талии нет и примерно с середины брюшко плавно сужается, на конце не такое заостренное, как у самца. Желтые боковые пятна на брюшке более обширные и при виде сверху черная дорсальная полоса значительно уже, чем у самца. Верхние придатки раздвинуты в основании, часто параллельны друг другу.
У неполовозрелых особей глаза коричневые; брюшко ярче, чем у половозрелой взрослой особи, с более обширным охристым и неравномерным черным рисунком на вершине сегментов III-X, как у неполовозрелой Libellula fulva; на крыльях широко распространен золотистый оттенок.
Несколько сходен с двумя более мелкими видами Libellula fulva и L. quadrimaculata. Окраска брюшка у самки или неполовозрелого самца L. fulva похожа на окраску E. bimaculata, но отличается темной линией у основания переднего крыла и темными вершинами крыльев. L. quadrimaculata у обоих полов имеет более или менее развитое черное пятно на узелках всех крыльев.
Длина 55-65 мм, брюшко 39-43 мм, заднее крыло 38-44 мм.
Лет: конец мая - начало июля.

♀ Фото © Владимир Бондарь, Быховский р-н, Могилевская обл.
Населяет крупные водоемы, небольшие озера или большие пруды, иногда рыбоводные, с площадью зеркала воды более 1 га и обширными открытыми водными поверхностями с хорошо развитыми растительными поясами осоки, тростника или камыша. В исключительных случаях заселяет медленно текущие водотоки (каналы и различные участки крупных медленных рек, где практически отсутствует течение). Плавающие и надводные слои гидрофитов, состоящие из Nymphea alba, Nuphar lutea, Polygonum amphibium, Myriophyllum spicatum, Potamogeton spp. важны для откладывания яиц и развития личинок. Конфигурация берегов может быть изменчивая, иногда пологая или более крутая. Наличие сбалансированной ихтиофауны, «рационально» управляемой (рыболовный пруд с устойчивым зарыблением), не препятствует развитию вида.
Для многих мест обитания характерна лесная или полулесная среда рядом с водоемами, которая, вероятно, играет роль во время созревания, отдыха и охоты. За исключением самок, ищущих субстрат для кладки яиц, наблюдение взрослых особей, отдыхающих на растительности на берегу, является исключением. Кормятся вокруг деревьев, на кронах которых отдыхают сидя вертикально. Максимальная продолжительность жизни взрослой особи оценивается в 30 дней.
Обитает совместно с Erythromma najas, Cordulia aenea, Somatochlora metallica и Orthetrum cancellatum. В некоторых местах регулярно развивается с Aeshna grandis.
Для многих мест обитания характерна лесная или полулесная среда рядом с водоемами, которая, вероятно, играет роль во время созревания, отдыха и охоты. За исключением самок, ищущих субстрат для кладки яиц, наблюдение взрослых особей, отдыхающих на растительности на берегу, является исключением. Кормятся вокруг деревьев, на кронах которых отдыхают сидя вертикально. Максимальная продолжительность жизни взрослой особи оценивается в 30 дней.
Обитает совместно с Erythromma najas, Cordulia aenea, Somatochlora metallica и Orthetrum cancellatum. В некоторых местах регулярно развивается с Aeshna grandis.

© Владимир Бондарь, Быховский р-н, Могилевская обл.
Информации о плотности популяции очень мало. Оценить эти данные тем труднее, что численность популяций сильно различается по годам и между стациями. В некоторых местностях лишь изредка обнаруживается единицы экзувиев, в то время как в других обнаруживается несколько сотен за утро исследования. Кроме того, на этой же стации можно наблюдать массовое появление один год (более 1000 экзувиев), тогда как в предыдущем году их было обнаружено менее 10. Взрослые особи наблюдаются редко, даже если на участке обнаружено несколько сотен экзувиев: поэтому обнаружение популяций и оценка численности в основном основаны на экзувиях. Обнаружение самцов, патрулирующих посреди водоемов, может лишь позволить обнаружить популяцию, но не оценить ее численность.
Поскольку взрослых наблюдают редко, мало что известно об их поведении. Их перемещения вне водоемов, по-видимому, никогда не изучались. После вылета они удаляются от мест размножения, что объясняет немногочисленность имеющихся наблюдений. Они также могут оставаться скрытыми среди лесной растительности вокруг занятых водоемов. По мере созревания стрекозы время от времени и ненадолго возвращаются к местам откладки яиц. На этом этапе они также остаются незаметными на месте размножения, так как летают над водой, вдали от берегов. Самки, видимо, ищут места для откладывания яиц в основном утром или с наступлением темноты. Также сообщалось о наблюдениях самцов в полете утром или вечером во время заката, в то время, когда большинство других стрекоз неактивны.
В целом известно, что Cordulidae удаляются от места своего появления, средний радиус передвижений составляет от 1 до 10 км. Вполне вероятно, что это относится и к данному виду. Взрослых часто можно увидеть охотящихся на лесных тропинках в нескольких километрах от мест размножения.
Поскольку взрослых наблюдают редко, мало что известно об их поведении. Их перемещения вне водоемов, по-видимому, никогда не изучались. После вылета они удаляются от мест размножения, что объясняет немногочисленность имеющихся наблюдений. Они также могут оставаться скрытыми среди лесной растительности вокруг занятых водоемов. По мере созревания стрекозы время от времени и ненадолго возвращаются к местам откладки яиц. На этом этапе они также остаются незаметными на месте размножения, так как летают над водой, вдали от берегов. Самки, видимо, ищут места для откладывания яиц в основном утром или с наступлением темноты. Также сообщалось о наблюдениях самцов в полете утром или вечером во время заката, в то время, когда большинство других стрекоз неактивны.
В целом известно, что Cordulidae удаляются от места своего появления, средний радиус передвижений составляет от 1 до 10 км. Вполне вероятно, что это относится и к данному виду. Взрослых часто можно увидеть охотящихся на лесных тропинках в нескольких километрах от мест размножения.

Фото © Семён Левый, Борисовский р-н, Минская обл.
Первые взрослые особи появляются в местах размножения после периода созревания, длящегося около 10 дней (хотя по другим данным, созревание длится 2-3 недели). У водоема в репродуктивный период имаго проводят относительно мало времени, особенно самки. Это, а также отличные летные способности, приводят к тому, что этот вид практически не наблюдается и еще реже отлавливается: только 30% данных касаются наблюдения за активными имаго, 60% - находки экзувий и 10% - отлов личинок или наблюдения за отрождениями.
Патрульный полет самцов длительный, проходит в нескольких метрах от берега над открытой водой на небольшой высоте (около 0,5 м). Самцы обустраивают и защищают четко определенные, но никогда не очень большие территории (от 10 до 30 м), которые почти всегда находятся далеко от берега над открытой водой. Их патрульный полет довольно медленный и устойчивый, часто с линейной траекторией, образующей большие петли примерно в 30-60 см от поверхности воды. Если другой представитель Anisoptera приближается к его территории, самец может быть очень агрессивным. Он внезапно ускоряется, преследуя нарушителя в погоне, которая может увести его далеко от его территории (более 50 м в высоту и 100 м вдаль). Таким образом, период отсутствия самца до возвращения на свою территорию может быть как достаточно коротким (несколько минут), так и очень длительным (более двух часов).
Патрульный полет самцов длительный, проходит в нескольких метрах от берега над открытой водой на небольшой высоте (около 0,5 м). Самцы обустраивают и защищают четко определенные, но никогда не очень большие территории (от 10 до 30 м), которые почти всегда находятся далеко от берега над открытой водой. Их патрульный полет довольно медленный и устойчивый, часто с линейной траекторией, образующей большие петли примерно в 30-60 см от поверхности воды. Если другой представитель Anisoptera приближается к его территории, самец может быть очень агрессивным. Он внезапно ускоряется, преследуя нарушителя в погоне, которая может увести его далеко от его территории (более 50 м в высоту и 100 м вдаль). Таким образом, период отсутствия самца до возвращения на свою территорию может быть как достаточно коротким (несколько минут), так и очень длительным (более двух часов).

♀ Фото © Владимир Бондарь, Быховский р-н, Могилевская обл.
Спаривание чаще всего происходит в пологе леса или среди тростниковых зарослей, окаймляющих водоем.
Самка откладывает яйца в одиночку на лету, при этом ударяет острием брюшка по поверхности воды, как бы выбивая яйца из яйцеклада. Яйца откладываются в воду (иногда в нескольких метрах от берега) или на водные растения, их оболочки разбухают, и кладки приобретают вид своеобразных студенистых (слизистых) шнуров, достигающих в длину 30-50 см и в ширину 1-2 см. Один конец шнура обыкновенно прикрепляется к растению (Potamogeton, Ceratophyllum и мн. др.) и шнуры плавают параллельно поверхности воды. Каждый шнур содержит по несколько сотен яиц, размещенных по периферии шнура. Имеются указания, что самка откладывает около 2000 яиц. Яйца продолговатые, желто-коричневые, длиной 7,5 мм, шириной 4 мм.
Яйца развиваются очень короткое время (примерно 3 недели); из них вылупляется предличинка, которая тут же линяет на личиночку. Последняя длиной 2 мм, шириной 1,5 мм, совершенно прозрачная, за исключением головы, переднегруди и заднего конца тела; через тонкую прозрачную кожу просвечивают 2 ствола боковых трахей. Голова личиночки широкая, большая; антенны длинные, 3-члениковые; глаза маленькие; на затылке расположены 2 длинных шипа в виде 2 рожков, слабо искривленных на вершине.
Самка откладывает яйца в одиночку на лету, при этом ударяет острием брюшка по поверхности воды, как бы выбивая яйца из яйцеклада. Яйца откладываются в воду (иногда в нескольких метрах от берега) или на водные растения, их оболочки разбухают, и кладки приобретают вид своеобразных студенистых (слизистых) шнуров, достигающих в длину 30-50 см и в ширину 1-2 см. Один конец шнура обыкновенно прикрепляется к растению (Potamogeton, Ceratophyllum и мн. др.) и шнуры плавают параллельно поверхности воды. Каждый шнур содержит по несколько сотен яиц, размещенных по периферии шнура. Имеются указания, что самка откладывает около 2000 яиц. Яйца продолговатые, желто-коричневые, длиной 7,5 мм, шириной 4 мм.
Яйца развиваются очень короткое время (примерно 3 недели); из них вылупляется предличинка, которая тут же линяет на личиночку. Последняя длиной 2 мм, шириной 1,5 мм, совершенно прозрачная, за исключением головы, переднегруди и заднего конца тела; через тонкую прозрачную кожу просвечивают 2 ствола боковых трахей. Голова личиночки широкая, большая; антенны длинные, 3-члениковые; глаза маленькие; на затылке расположены 2 длинных шипа в виде 2 рожков, слабо искривленных на вершине.

♂ Фото © Семён Левый, г. Минск
Нимфа. Личинки серо-коричневые, с широким удлиненным слабо волосатым телом. Голова небольшая, широкая, назади узкая, почти в 2 раза уже ширины; лоб слегка выпуклый, коричневый, с темными пятнами, посредине с белой полосой, разветвляющейся у основания и окружающей слабо заметные глазки; темя отделено от затылка волнистой линией, маленькое; глаза выпуклые, конические, округлые на вершине; антенны длинные, заходят за дистальный край боковой лопасти, покрыты тонкими волосками, первые два членика толстые, темные, остальные тонкие, светлые; затылок коричневый, с более светлой гладкой широкой продольной полосой посредине, по обе стороны которой находятся 2 большие светлые гладкие широкие полосы (полосы слабо различимы у личинок последней стадии и хорошо видимы у личинок более ранних стадий); посредине последние две полосы слабо усеяны щетинками и несут по одному выдающемуся коническому островершинному покрытому щетинками шипу; внешние стороны затылка прямые, усеянные маленькими щетинками, задние углы округлые, задний край слабо вогнутый; вся поверхность затылка покрыта маленькими шипами. Переднегрудь такой же ширины, как и задний край затылка; задняя сторона округлая, со вздутым краем; внешние стороны слегка приподнятые; средне- и заднегрудь дорсально коричневые, вентрально светлые. Крыловые чехлы достигают VI сегмента. Ноги очень длинные, бедра последней пары доходят до VIII сегмента, цилиндрические, тонкие, широко расставленные у основания; ноги передней пары более сближены; бедра изогнутые; голени прямые, на задней ноге несколько длиннее бедер; бедра и голени с 2 темными кольцами и усеяны темными мелкими пятнами. Брюшко широкое, дорсально выпуклое, вентрально плоское, укороченное, длина менее чем в 2 раза превосходит ширину; наибольшая ширина брюшка в области VII и VIII сегментов, начиная от VII сегмента постепенно суживается к усеченному заднему концу; дорсально с темными пятнами и широкими светлыми отметинами; по длине все сегменты, за исключением Х, более или менее равны между собой; X сегмент очень короткий и помещается в выемке заднего края IX сегмента; дорсальные шипы на (II) III-IX сегментах, первый шип тонкий, острый, конический, остальные более широкие, с боков приплюснутые, с острой изогнутой назад вершиной, на IX сегменте шип искривлен сильнее, чем на предшествующем; латеральные шипы на VIII и IX сегментах острые, изогнутые; на VIII сегменте шип короткий, равен 1/4 длины края сегмента; на IX сегменте шип очень длинный и сильно искривленный, заходящий за анальную пирамиду. Анальная пирамида такой же длины, как IX и X сегменты, взятые вместе, относительно маленькая, треугольная, прямая, снабженная тонкими волосками; дорсальный придаток короче церков на 1/4, с острой вершиной и сильно расширенным основанием; основная часть дорсального придатка самца равна половине его длины, широкая; церки длинные, островершинные; церкоиды конические, с острой короткой вершиной, немного короче дорсального придатка. Нимфы последних возрастов размером от 28 до 32 мм.

© Оксана С., Минский р-н
Личинки являются обитателями стоячих водоемов, главным образом водоемов с большой поверхностью воды; в реках живут в тихих больших заводях, где в прибрежной зоне создаются условия, подходящие к таковым в стоячих водоемах. Вода может быть от кислой до относительно щелочной. Вид встречается, в частности, в торфяных прудах, покрытых сфагновым мхом, а также в щелочных озерах. Личинки ведут придонный образ жизни. Обычно держатся на мелководье (менее 1 м), но иногда могут быть найдены на глубине более 2 м. У них суточный цикл, днем они остаются на дне, а ночью становятся активными в поисках пищи (особенно среди водной растительности). Личинки раннего возраста также могут быть очень подвижными в течение дня.
В своем развитии используют разные участки, перемещаясь между плавающими зарослями растительности и илистым дном водоемов. Для зимовки зарываются в субстрат. Заканчивают развитие за 2-3, иногда за 4 года.
Вылупление взрослых стрекоз, как правило, носит массовый характер, хотя может растягиваться на 2-3 недели. Отрождение происходит в основном в первых двух метрах от берега и на высоте менее 50 см, но личинки могут использовать в качестве субстрата и деревья. Их можно встретить далеко от берега, иногда до ста метров, особенно в зарослях ежевики и крапивы, или на деревьях, иногда на высоте более 10 м. Некоторые личинки могут пересекать тропы или даже дороги, чтобы выйти за их пределы, если структура растительности на берегу неблагоприятна.
Обычно только что вылупившиеся тенеральные особи быстро летят к верхушкам деревьев и удаляются от места своего появления, даже не исследуя его. Созревающие молодые особи исчезают из мест своего появления и часто не возвращаются в эти места в период размножения. Точно так же имаго в репродуктивной фазе лишь изредка наблюдаются в местах вылета, возможно, они посещают другие места, которые кажутся им благоприятными. Расстояние расселения этого вида неизвестно. Можно предположить, что особи способны перемещаться на несколько десятков километров, но ничто не позволяет утверждать, что такие расстояния преодолеваются часто.
В своем развитии используют разные участки, перемещаясь между плавающими зарослями растительности и илистым дном водоемов. Для зимовки зарываются в субстрат. Заканчивают развитие за 2-3, иногда за 4 года.
Вылупление взрослых стрекоз, как правило, носит массовый характер, хотя может растягиваться на 2-3 недели. Отрождение происходит в основном в первых двух метрах от берега и на высоте менее 50 см, но личинки могут использовать в качестве субстрата и деревья. Их можно встретить далеко от берега, иногда до ста метров, особенно в зарослях ежевики и крапивы, или на деревьях, иногда на высоте более 10 м. Некоторые личинки могут пересекать тропы или даже дороги, чтобы выйти за их пределы, если структура растительности на берегу неблагоприятна.
Обычно только что вылупившиеся тенеральные особи быстро летят к верхушкам деревьев и удаляются от места своего появления, даже не исследуя его. Созревающие молодые особи исчезают из мест своего появления и часто не возвращаются в эти места в период размножения. Точно так же имаго в репродуктивной фазе лишь изредка наблюдаются в местах вылета, возможно, они посещают другие места, которые кажутся им благоприятными. Расстояние расселения этого вида неизвестно. Можно предположить, что особи способны перемещаться на несколько десятков километров, но ничто не позволяет утверждать, что такие расстояния преодолеваются часто.

♀ Фото © Семён Левый, Борисовский р-н, Минская обл.

Photo © pavelbykau / iNaturalist.org. Воложинский р-н, Минская обл. CC BY-NC 4.0

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
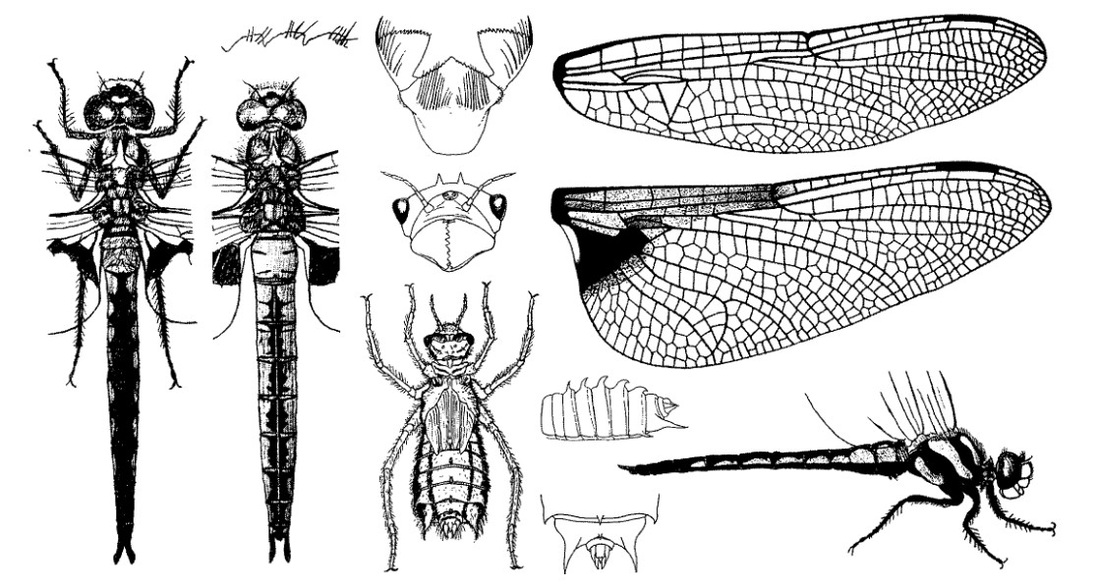
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Smallshire D., Swash A. "Eurasian Baskettail. Epitheca bimaculata" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 250-251.
8. Monnerat C. "Epitheca bimaculata (Charpentier,1825) Cordulie à deux taches - Zweifleck"/ dans: Wildennuth H., Gonseth Y., Maibach A. (éds.) Odonata. Les Libellules de Suisse. Fauna Helvetica 11. Centre suisse de cartographie de la faune Schweizerische Entornologische Gesellschaft, 2005. pp. 265-269.
9. Monnerat C. "Fiches de protection espèces - Libellules - Epitheca bimaculata". Groupe de travail pour la conservation des Libellules de Suisse GTCLS - Bâle, CSCF info fauna - Neuchâtel, OFEV, Berne, 2013. 5 p.
10. Merlet F., Houard X. "Synthèse bibliographique sur les traits de vie de l’Épithèque bimaculée (Epitheca bimaculata (Charpentier, 1825)) relatifs а ses déplacements et а ses besoins de continuités écologiques". Office pour les insectes et leur environnement & Service du patrimoine naturel du Muséum national d’Histoire naturelle. Paris, 2012. - 8 pp.
11. Mauersberger R., Beutler H. "Zweifleck – Epitheca bimaculata (Charpentier 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 100-101.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Smallshire D., Swash A. "Eurasian Baskettail. Epitheca bimaculata" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 250-251.
8. Monnerat C. "Epitheca bimaculata (Charpentier,1825) Cordulie à deux taches - Zweifleck"/ dans: Wildennuth H., Gonseth Y., Maibach A. (éds.) Odonata. Les Libellules de Suisse. Fauna Helvetica 11. Centre suisse de cartographie de la faune Schweizerische Entornologische Gesellschaft, 2005. pp. 265-269.
9. Monnerat C. "Fiches de protection espèces - Libellules - Epitheca bimaculata". Groupe de travail pour la conservation des Libellules de Suisse GTCLS - Bâle, CSCF info fauna - Neuchâtel, OFEV, Berne, 2013. 5 p.
10. Merlet F., Houard X. "Synthèse bibliographique sur les traits de vie de l’Épithèque bimaculée (Epitheca bimaculata (Charpentier, 1825)) relatifs а ses déplacements et а ses besoins de continuités écologiques". Office pour les insectes et leur environnement & Service du patrimoine naturel du Muséum national d’Histoire naturelle. Paris, 2012. - 8 pp.
11. Mauersberger R., Beutler H. "Zweifleck – Epitheca bimaculata (Charpentier 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 100-101.
