СТРЕЛКА КОПЬЕНОСНАЯ
Coenagrion hastulatum Charpentier, 1825

♂ Фото © Николай Гурков, Витебский р-н
Coenagrion hastulatum Charpentier, 1825 (стрелка копьеносная)
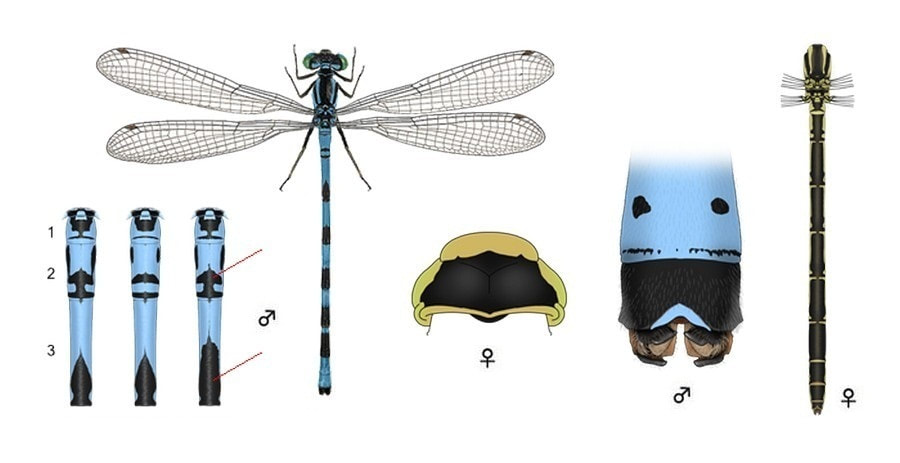
Широкая голова, сверху вдвое шире своей длины. У обоих полов имеется светлая поперечная полоса между затылочными пятнами. Глаза и «лицо» снизу ярко-зеленые, сверху глаза темные или черные. Задний край переднеспинки треугольный, посредине вытянут назад, полностью или частично светлый, образует посредине тупой угол, без выступа. У обоих полов на боках груди есть так называемая «шпора». Крылья прозрачные. Птеростигма узкая (равна 1 ячейке), одноцветная. Ноги черные или темно-серые.
♂: Общая окраска голубая или голубовато-зеленая. Бока груди бледно-зеленовато-голубые. Задний край переднеспинки треугольный, голубой. Брюшко с черным рисунком на I-VII сегментах, VIII-IX голубые, с переменными черными метками на сегменте IX (обычно двумя маленькими черными пятнами) и, у некоторых, на VIII. Сегмент X - дорсально полностью черный, но латерально и вентрально - голубой. Рисунок II тергита состоит обычно из трех темных пятен, из которых среднее T-образное и касается заднего края тергита (не всегда), а два других в виде полосок расположены по бокам впереди; редко боковые полоски сливаются со средним пятном. Сегмент III имеет черную метку в виде «наконечника копья». Нижние анальные придатки длиннее верхних и сверху образуют короткие загнутые «хоккейные клюшки».
♀: Голубовато- или зеленовато-желтая (молодая розоватая), брюшко с черным рисунком, с широкими бронзово-черными пятнами на дорсальной поверхности брюшка. Задний край переднеспинки треугольный, обрамленный сзади желтовато-зеленой линией. Тергит II с черный от переднего до заднего конца членика; тергиты III-X сверху черные с узким светлым пояском у переднего края. VIII тергит сплошь черный.
♂: Общая окраска голубая или голубовато-зеленая. Бока груди бледно-зеленовато-голубые. Задний край переднеспинки треугольный, голубой. Брюшко с черным рисунком на I-VII сегментах, VIII-IX голубые, с переменными черными метками на сегменте IX (обычно двумя маленькими черными пятнами) и, у некоторых, на VIII. Сегмент X - дорсально полностью черный, но латерально и вентрально - голубой. Рисунок II тергита состоит обычно из трех темных пятен, из которых среднее T-образное и касается заднего края тергита (не всегда), а два других в виде полосок расположены по бокам впереди; редко боковые полоски сливаются со средним пятном. Сегмент III имеет черную метку в виде «наконечника копья». Нижние анальные придатки длиннее верхних и сверху образуют короткие загнутые «хоккейные клюшки».
♀: Голубовато- или зеленовато-желтая (молодая розоватая), брюшко с черным рисунком, с широкими бронзово-черными пятнами на дорсальной поверхности брюшка. Задний край переднеспинки треугольный, обрамленный сзади желтовато-зеленой линией. Тергит II с черный от переднего до заднего конца членика; тергиты III-X сверху черные с узким светлым пояском у переднего края. VIII тергит сплошь черный.

♂ Фото © Виталий Сирота, г. Минск
Бледно-зеленовато-голубые бока могут внешне напоминать C. lunulatum, хотя эти два вида различаются степенью голубизны на брюшке, которая более обширна у C. hastulatum. Самцы обоих видов обычно имеют короткую черную полосу по обе стороны от сегмента II брюшка, но они обычно различаются по форме центральной отметки на этом сегменте, которая обычно имеет стреловидную форму у C. hastulatum. Однако эта отметка изменчива и может быть редуцирована до пятна в виде продольной полоски, как у Enallagma cyathigerum, или до поперечной полосы, как у C. lunulatum. Последний вид также отличается от С. hastulatum формой заднего края переднеспинки и анальных придатков.
Самки темные, с зелеными боками груди и брюшка, внешне напоминают самку E. cyathigerum, с которой они иногда сосуществуют, но лишены генитального шипа на вентральной поверхности VIII сегмента брюшка.
Длина 31-34 мм, брюшко 28-30 мм, заднее крыло 17-22 мм.
Лет: середина мая - август.
Самки темные, с зелеными боками груди и брюшка, внешне напоминают самку E. cyathigerum, с которой они иногда сосуществуют, но лишены генитального шипа на вентральной поверхности VIII сегмента брюшка.
Длина 31-34 мм, брюшко 28-30 мм, заднее крыло 17-22 мм.
Лет: середина мая - август.

♂ Фото © Николай Гурков, Витебский р-н
Населяет стоячие водоемы любого типа. Размножается в защищенных и заболоченных водоемах с обильной надводной растительностью и с оптимальной глубиной около 50 см. Также размножается на неглубоких защищенных окраинах озер среднего размера, где медленно летает среди высокого тростника и осоки, часто садясь на эти растения. Излюбленные участки обычно мезотрофны и часто содержат кувшинку белую, вахту трехлистную и узколистные осоки, а также небольшие краевые сфагновые участки с болотным миртом и пушицей обыкновенной.
В целом, считается, что требования к среде обитания до сих пор недостаточно изучены. Предпочтение болотных водоемов очевидно, однако старые песчаные или гравийные карьеры, неглубокие пруды или даже канавы всегда являются частью спектра их среды обитания, при условии, что они имеют низкий уровень питательных веществ в сочетании с разнообразной структурой растительности, которая не слишком высока.
В кислых болотных озерах с преобладанием торфяных мхов встречается довольно редко; основное внимание, вероятно, уделяется субнейтральным небольшим водоемам с затопленными растительными структурами. По-видимому, все репродуктивные водоемы находятся в лесу или, по крайней мере, вдали от интенсивно используемых сельскохозяйственных ландшафтов. Эти воды всегда имеют очень малые площади водосбора и почти никогда не проточные.
Вид может переносить временное высыхание среды обитания.
Предпочтительными местами обитания и охоты часто являются влажные луга и напочвенная растительность открытых участков в прилегающем лесу, например, на тропах и полянах. Обнаружены значительные изменения местоположения, в том числе между различными водоемами.
Оценка продолжительности жизни в имагинальной фазе примерно в 25-30 дней. Максимальная продолжительность жизни значительно выше, потому что в Германии двух самцов обнаружили через 32 дня после их маркировки, что добавляет им еще минимум 5 дней фазы созревания.
В целом, считается, что требования к среде обитания до сих пор недостаточно изучены. Предпочтение болотных водоемов очевидно, однако старые песчаные или гравийные карьеры, неглубокие пруды или даже канавы всегда являются частью спектра их среды обитания, при условии, что они имеют низкий уровень питательных веществ в сочетании с разнообразной структурой растительности, которая не слишком высока.
В кислых болотных озерах с преобладанием торфяных мхов встречается довольно редко; основное внимание, вероятно, уделяется субнейтральным небольшим водоемам с затопленными растительными структурами. По-видимому, все репродуктивные водоемы находятся в лесу или, по крайней мере, вдали от интенсивно используемых сельскохозяйственных ландшафтов. Эти воды всегда имеют очень малые площади водосбора и почти никогда не проточные.
Вид может переносить временное высыхание среды обитания.
Предпочтительными местами обитания и охоты часто являются влажные луга и напочвенная растительность открытых участков в прилегающем лесу, например, на тропах и полянах. Обнаружены значительные изменения местоположения, в том числе между различными водоемами.
Оценка продолжительности жизни в имагинальной фазе примерно в 25-30 дней. Максимальная продолжительность жизни значительно выше, потому что в Германии двух самцов обнаружили через 32 дня после их маркировки, что добавляет им еще минимум 5 дней фазы созревания.

© Виталий Сирота, г. Минск
Летает довольно слабо, редко вылетает на открытую воду; отдает предпочтение зарослям осоки, обычно располагаясь внизу, близко к поверхности воды. Часто обитает совместно с E. cyathigerum, а также может быть замечен с C. puella, Lestes sponsa, Pyrrhosoma nymphula, Libellula quadrimaculata и другими видами.
Территориальное поведение не зарегистрировано, и самцы могут сосуществовать при высокой плотности популяции. При солнечной погоде самцы низко летают над затопленными структурированными водоемами или пересекают заиленные участки с мелкой осокой, сабельником (Potentilla palustris) и отдельными стеблями тростника. Самки наблюдаются редко, отчасти потому, что их трудно отличить от самок C. puella, которые также очень распространены в типичной среде обитания.
Спаривание происходит как в прибрежной зоне, так и в кормовых местообитаниях. При отсутствии беспокойства копуляция в форме кольца продолжается до 30 минут.
Самки, откладывающие яйца, при приближении чужих самцов, защищаются, поднимая брюшко. Яйца в основном откладываются в тандеме. Самка откладывает их в погруженные побеги, чаще в черешки листьев рдеста, при этом она часто полностью погружается в воду, а сопровождающий ее самец спускается только до половины. Хотя по другим данным, оба партнера часто вместе уходят под воду. Такие погружения длятся от 6 до 23 минут.
Эмбриональное развитие короткое, личинки обычно вылупляются через 2-3 недели после откладки яиц.
Территориальное поведение не зарегистрировано, и самцы могут сосуществовать при высокой плотности популяции. При солнечной погоде самцы низко летают над затопленными структурированными водоемами или пересекают заиленные участки с мелкой осокой, сабельником (Potentilla palustris) и отдельными стеблями тростника. Самки наблюдаются редко, отчасти потому, что их трудно отличить от самок C. puella, которые также очень распространены в типичной среде обитания.
Спаривание происходит как в прибрежной зоне, так и в кормовых местообитаниях. При отсутствии беспокойства копуляция в форме кольца продолжается до 30 минут.
Самки, откладывающие яйца, при приближении чужих самцов, защищаются, поднимая брюшко. Яйца в основном откладываются в тандеме. Самка откладывает их в погруженные побеги, чаще в черешки листьев рдеста, при этом она часто полностью погружается в воду, а сопровождающий ее самец спускается только до половины. Хотя по другим данным, оба партнера часто вместе уходят под воду. Такие погружения длятся от 6 до 23 минут.
Эмбриональное развитие короткое, личинки обычно вылупляются через 2-3 недели после откладки яиц.

© Николай Гурков, Витебский р-н
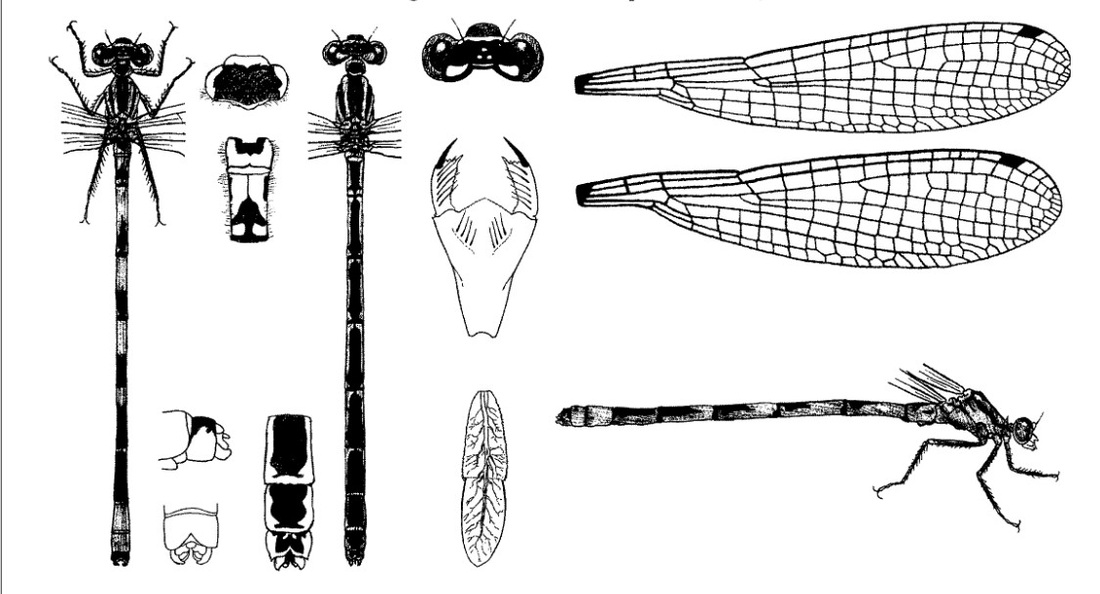
Нимфа. Окраска тела варьирует: зеленоватая, желтоватая или коричневатая. Тело гладкое, покрыто маленькими темно-коричневыми хитиновыми точками. Длина 21-23 мм. Голова широкая (3,5-4 мм), почти в 2 раза шире длины, округлая впереди, усеяна темными точками, варьирующими в размерах; глаза средних размеров, относительно выпуклые, расположены спереди по сторонам головы; антенны с более темною базальною частью; затылок с сильно скошенными внешними сторонами, с сильно округлыми задними углами, покрытыми довольно большими шипами, с глубоко вогнутым задним краем; под кожей затылка в центре между глаз просвечивают 2 трахейных ствола в виде темных полос. Маска в состоянии покоя доходит до основания средних ног; подбородок у дистального края в 2 раза шире, чем у основания; передний край средней лопасти выдается вперед в виде тупого угла и тонко зазубрен; внешний край боковых лопастей маски выпуклый; внутренний край слабо вогнутый, зазубренный; дистальный край приблизительно с 6 зубчиками; конечный крючок резко выраженный, клювообразный; подвижный крючок довольно длинный, изогнутый, острый. Переднегрудь вдвое уже головы, значительно шире своей длины, с тупоугольными темными внешними краями, задним выпуклым краем, с 2 темными продольными полосами, продолжающимися на голову. Крыловые чехлы заходят на IV сегмент. Бедра всех пар ног с темным кольцом близ дистального конца, немного короче и толще голеней и слабо изогнутые; бедра и голени вооружены шипами, а лапки щетинками. Брюшко цилиндрическое, постепенно суживающееся к вершине, дорсально с тонкой светлой продольной срединной полосой и со светлыми внешними краями; светлые полосы без шипиков; межсегментальные пространства с чередующимися светлыми и темными пятнами; острые внешние края несут ряд шипиков, из которых самый последний наибольших размеров; задний край Х сегмента обрамлен шипами; вентральная сторона светлее дорсальной. Хвостовые жаберные пластинки ланцетовидной формы; длина их приблизительно в 5 раз превосходит ширину; с более или менее ясным заострением на вершине; средняя пластинка значительно шире и более закругленная на вершине, чем боковые; главная трахея в средней пластинке проходит только в базальной половине, а в боковых она простирается и в дистальную часть; светлый тонкий поперечный узелковый шов идет приблизительно по середине пластинок или немного дистальнее, образуя прямые углы с главным трахейным стволом и деля пластинки на 2 части; узелки сильные, четкие; пластинки без пятен или со слабым поперечным рисунком; разветвление трахей густое и четкое. Створки яйцеклада самки доходят или почти доходят до конца Х стернита и заканчиваются 2 направленными вниз остриями, на нижнем крае усеяны рядом довольно больших шипов. Субанальные пластинки самки узкие, на вершинах заостренные; у самца они широкие, на вершинах округлые.

© Николай Гурков, Витебский р-н
Личинки заселяют самые различные водоемы: озера, пруды, болота, лужи, канавы, ручьи, заводи медленно текущих небольших речек, но предпочитают небольшие пруды, болота, лужи, богатые водной растительностью. Живут обычно на глубине 0,1-0,5-1,0 м среди водных растений, особенно растущих на толстом слое торфа. Личинки очень подвижны, скрываются при малейшем беспокойстве, однако ищут себе пищу днем и ночью. В зависимости от ситуации могут как охотиться из засады, так и незаметно приближаться к своей добыче. Наиболее частыми жертвами являются ветвистоусые рачки (Cladocera) и веслоногие рачки рода Cyclops. При появлении крупных личинок Aeshna spp., которых считают опасными, замирают и остаются неподвижными.
Развитие личинок проходит 10-13 стадий и, вероятно, занимает 1 или 2 года. Данные очень различны, например, для южной Швеции указывается 1 год, для равнин Швейцарии более 1 года, а для горных регионов 2-3 года. Для европейской части бывшего СССР - 1-2 года. Вероятно, сроки связаны не только с географической широтой и высотой над уровнем моря, а и с конкретными условиями водоемов. В небольших водоемах, которые могут пересыхать, личинкам удается некоторое время выживать, зарываясь во влажный торф, однако их развитие затормаживается.
В теплые годы период отрождения начинается в конце апреля, в холодные - несколько позже. Большинство имаго вылупляются между серединой мая и серединой июня, период отрождения заканчивается в первой половине июля. Согласно разным исследованиям в разные годы фаза отрождения и максимум отрождений различны. Например, фаза отрождения 29 дней и максимум наступил на 5-й день, либо 24 дня и максимум на 10-й день. Видимо, сроки зависят как от условий водоема, так и от погоды в конкретный год.
Для отрождения личинки вылазят из воды в основном утром, поднимаются на несколько сантиметров над ее поверхностью на вертикальные части растений. Сразу после отрождения имаго покидают водоем. Данные о тенеральных особях несколько разнятся и, видимо, зависят от конкретных условий. По одним данным, они далеко не отлетают и держатся у водоема, по другим, - могут отлетать от водоема во влажные леса.
Развитие личинок проходит 10-13 стадий и, вероятно, занимает 1 или 2 года. Данные очень различны, например, для южной Швеции указывается 1 год, для равнин Швейцарии более 1 года, а для горных регионов 2-3 года. Для европейской части бывшего СССР - 1-2 года. Вероятно, сроки связаны не только с географической широтой и высотой над уровнем моря, а и с конкретными условиями водоемов. В небольших водоемах, которые могут пересыхать, личинкам удается некоторое время выживать, зарываясь во влажный торф, однако их развитие затормаживается.
В теплые годы период отрождения начинается в конце апреля, в холодные - несколько позже. Большинство имаго вылупляются между серединой мая и серединой июня, период отрождения заканчивается в первой половине июля. Согласно разным исследованиям в разные годы фаза отрождения и максимум отрождений различны. Например, фаза отрождения 29 дней и максимум наступил на 5-й день, либо 24 дня и максимум на 10-й день. Видимо, сроки зависят как от условий водоема, так и от погоды в конкретный год.
Для отрождения личинки вылазят из воды в основном утром, поднимаются на несколько сантиметров над ее поверхностью на вертикальные части растений. Сразу после отрождения имаго покидают водоем. Данные о тенеральных особях несколько разнятся и, видимо, зависят от конкретных условий. По одним данным, они далеко не отлетают и держатся у водоема, по другим, - могут отлетать от водоема во влажные леса.

♂ Фото © Николай Гурков, Витебский р-н
Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Spearhead Bluet. Coenagrion hastulatum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 74-75.
9. Smallshire D., Swash A. "Northern Damselfly. Coenagrion hastulatum" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 80-81.
10. Baumann K., Jödicke R., Pix A. "Coenagrion hastulatum – Speer-Azurjungfer" / Atlas der Libellen in Niedersachsen/Bremen. s. 84-89.
11. Küry D., Wildermuth H. "Fiches de protection espèces – Libellules – Coenagrion hastulatum" Groupe de travail pour la conservation des Libellules de Suisse, CSCF info fauna, Neuchâtel et Office fédéral de l’environnement, Berne, 2013. 5 p.
12. Mauersberger R. "Speer-Azurjungfer – Coenagrion hastulatum (Charpentier 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 36-37.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Spearhead Bluet. Coenagrion hastulatum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 74-75.
9. Smallshire D., Swash A. "Northern Damselfly. Coenagrion hastulatum" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 80-81.
10. Baumann K., Jödicke R., Pix A. "Coenagrion hastulatum – Speer-Azurjungfer" / Atlas der Libellen in Niedersachsen/Bremen. s. 84-89.
11. Küry D., Wildermuth H. "Fiches de protection espèces – Libellules – Coenagrion hastulatum" Groupe de travail pour la conservation des Libellules de Suisse, CSCF info fauna, Neuchâtel et Office fédéral de l’environnement, Berne, 2013. 5 p.
12. Mauersberger R. "Speer-Azurjungfer – Coenagrion hastulatum (Charpentier 1825)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 36-37.
