СТРЕКОЗА РЫЖАЯ
Libellula fulva Muller, 1764

♂ Фото © Денис Китель
Libellula fulva Muller, 1764 (стрекоза рыжая)
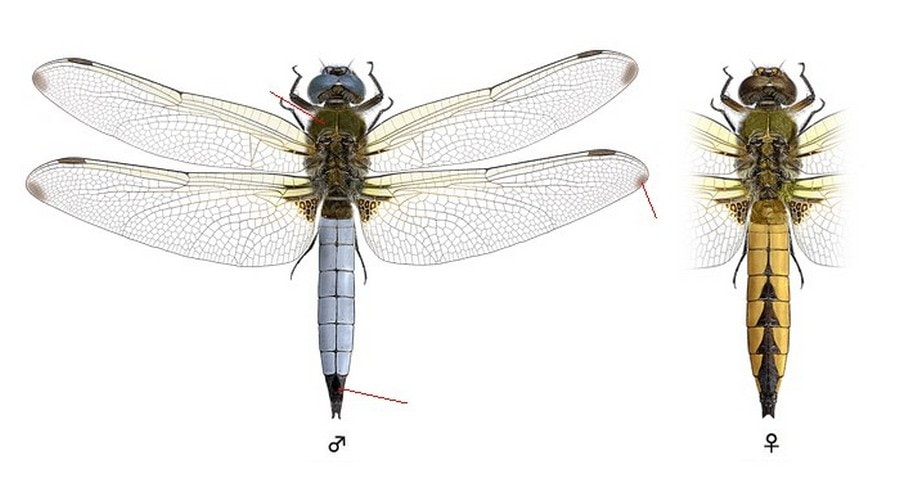
Нижняя губа вся черная. Грудь покрыта волосками, темная со светлыми предплечевыми полосами. Брюшко рыжеватое, широкое, особенно у самок, его средние сегменты вдвое шире своей длины.
Крылья с небольшими темными пятнами у основания, треугольные на заднем крыле и редуцированные до полосы на переднем крыле, с желтым оттенком спереди и часто с темными кончиками, особенно у самок. Птеростигма темно-серая или черноватая.
♂: Глаза голубовато-серые. Грудь черноватая. У взрослых брюшко покрыто голубоватым налетом. Он распространяется на III-VII сегменты брюшка, оставляя два передних и три задних сегмента черными, и скрывая срединную черную полоску. У основания и на VIII-X сегментах брюшко черноватое, или становится целиком темным (у старых). Посередине брюшка часто хорошо заметны темные «шрамы от спаривания» (налет, сцарапанный ногами самок). I сегмент брюшка без шипиков.
♀: Глаза коричневые или серо-коричневые. Грудь оливково-коричневая. Брюшко охристо-коричневое или насыщенно рыжее, с черной полосой вдоль вершин сегментов IV-X, расширяющейся к кончику в ряды своеобразных «колокольчиков». С возрастом брюшко темнеет, иногда покрывается налетом.
Крылья с желтым оттенком спереди.
У неполовозрелых особей глаза коричневые, ярко-коричневая грудь и оранжевое брюшко, похожее на самку; крылья с янтарным оттенком и желтыми жилками спереди. Молодые особи обычно сохраняют такую окраску в течение 10 дней.
Крылья с небольшими темными пятнами у основания, треугольные на заднем крыле и редуцированные до полосы на переднем крыле, с желтым оттенком спереди и часто с темными кончиками, особенно у самок. Птеростигма темно-серая или черноватая.
♂: Глаза голубовато-серые. Грудь черноватая. У взрослых брюшко покрыто голубоватым налетом. Он распространяется на III-VII сегменты брюшка, оставляя два передних и три задних сегмента черными, и скрывая срединную черную полоску. У основания и на VIII-X сегментах брюшко черноватое, или становится целиком темным (у старых). Посередине брюшка часто хорошо заметны темные «шрамы от спаривания» (налет, сцарапанный ногами самок). I сегмент брюшка без шипиков.
♀: Глаза коричневые или серо-коричневые. Грудь оливково-коричневая. Брюшко охристо-коричневое или насыщенно рыжее, с черной полосой вдоль вершин сегментов IV-X, расширяющейся к кончику в ряды своеобразных «колокольчиков». С возрастом брюшко темнеет, иногда покрывается налетом.
Крылья с желтым оттенком спереди.
У неполовозрелых особей глаза коричневые, ярко-коричневая грудь и оранжевое брюшко, похожее на самку; крылья с янтарным оттенком и желтыми жилками спереди. Молодые особи обычно сохраняют такую окраску в течение 10 дней.

♀ Фото © Виктор Рындевич, Россонский р-н, Витебская обл.
Черный кончик брюшка заметен в полете и помогает отличить от взрослых самцов Orthetrum coerulescens и от L. depressa (сегмент X у L. depressa темный, но настолько мал, что едва заметен в полете, а его брюшко намного шире, чем у L. fulva). У основания крыльев имеется темное черно-бурое пятно, сильно редуцированное в передних крыльях, а также на кончиках всех крыльев темные пятна, что отличает вид от O. coerulescens и O. cancellatum, у которых крылья прозрачные.
Лет: конец мая - август.
Длина 42-45 мм, брюшко 26-29 мм, заднее крыло 35-38 мм.
Лет: конец мая - август.
Длина 42-45 мм, брюшко 26-29 мм, заднее крыло 35-38 мм.

♂ Фото © Владимир Бондарь, Могилевский р-н
Населяет стоячие или медленно текущие водоемы различных типов и размеров, с богатой растительностью и почти обязательно окруженных поясом тростниковой или камышовой растительности. Развивается в стоячих и малопроточных, по крайней мере, частично солнечных, от олиготрофных до эвтрофных, многолетних водоемах с большей частью прозрачной, от нейтральной до щелочной водой и преимущественно выраженными зонами заиления осоковых или тростниковых зарослей. В небольших проточных водоемах это могут быть и небольшие заросли из травянистых пород. Как правило, заселяются заводи более крупных рек, естественные и малые озера и пруды, а также канавы и каналы с медленным течением. Относится к числу видов, обитающих в проточных озерах. Например, в Бранденбурге к северу от Берлина озера составляют более 80% местообитаний, но упоминаются и торфяные разрезы, канавы и ручьи, а южнее - это заводи, каналы и песчаные карьеры.
Встречается вместе с Platycnemis pennipes, Calopteryx splendens и Gomphus vulgatissimus на реках, а также с Coenagrion pulchellum и Brachytron pratense на лугах возле заболоченной местности.
Встречается вместе с Platycnemis pennipes, Calopteryx splendens и Gomphus vulgatissimus на реках, а также с Coenagrion pulchellum и Brachytron pratense на лугах возле заболоченной местности.

♂ Фото © Владимир Бондарь, Могилевский р-н
С последней декады мая в водоемах размножения появляются самцы; высокая численность иногда может наблюдаться уже в конце мая, в других случаях с июня до середины июля. В большинстве лет лёт заканчивается в конце июля или начале августа.
Территориальные самцы агрессивны, садятся на маргинальную растительность на высоте до 1 м над водой и перехватывают пролетающих незваных гостей или самок. В Британии самая высокая постоянная плотность составляет около 9 на 100 м уреза воды, когда самцы активны, но поздним вечером некоторые из них могут садиться в непосредственной близости друг от друга.
Наблюдается территориальность самцов как на внутривидовом, так и на межвидовом уровнях. В Чехии средний размер территории представлял собой круг с полудиаметром 2 м. Каждая территория защищалась от проникновения другого самца. Межвидовая территориальность наблюдалась преимущественно в отношении Orhretrum coerulescens. Взаимные нападения обоих видов самцов наблюдались только в ветреную погоду. Во время штиля общее пространство было разделено следующим образом: самцы O. coerulescens приземлялись на землю и добывали добычу на высоте 50 см над землей, а самцы L. fulva садились на тростник на высоте 1 м и выше. В ветреную погоду самцы L. fulva перемещались ниже, на территорию O. coerulescens, что и являлось причиной частых столкновений.
Территориальные самцы агрессивны, садятся на маргинальную растительность на высоте до 1 м над водой и перехватывают пролетающих незваных гостей или самок. В Британии самая высокая постоянная плотность составляет около 9 на 100 м уреза воды, когда самцы активны, но поздним вечером некоторые из них могут садиться в непосредственной близости друг от друга.
Наблюдается территориальность самцов как на внутривидовом, так и на межвидовом уровнях. В Чехии средний размер территории представлял собой круг с полудиаметром 2 м. Каждая территория защищалась от проникновения другого самца. Межвидовая территориальность наблюдалась преимущественно в отношении Orhretrum coerulescens. Взаимные нападения обоих видов самцов наблюдались только в ветреную погоду. Во время штиля общее пространство было разделено следующим образом: самцы O. coerulescens приземлялись на землю и добывали добычу на высоте 50 см над землей, а самцы L. fulva садились на тростник на высоте 1 м и выше. В ветреную погоду самцы L. fulva перемещались ниже, на территорию O. coerulescens, что и являлось причиной частых столкновений.

♂ Фото © Татьяна Алешка, Вилейский р-н, Минская обл.
Самцы в основном наиболее активны с 11 до 17 часов. Самки наблюдаются у водоема только во время совокупления или яйцекладки, чаще после 14 часов.
Отмечена высокая чувствительность к погодным условиям. Взрослые не проявляют активности в пасмурную погоду. В малооблачные дни имаго были очень чувствительны к изменению яркости солнечного света. В ясные дни самцы наиболее активны с 11 до 16 часов, а в дни с дневными грозами имаго либо наблюдались ранним вечером, либо не появлялись вовсе.
Самцы находили места для отдыха в тростнике на высоте 1 м и выше. Когда они замечали добычу, устремлялись к ней, пытаясь поймать. Успешными или неуспешными были их попытки, они всегда возвращались на одну и ту же присаду.
Отмечена высокая чувствительность к погодным условиям. Взрослые не проявляют активности в пасмурную погоду. В малооблачные дни имаго были очень чувствительны к изменению яркости солнечного света. В ясные дни самцы наиболее активны с 11 до 16 часов, а в дни с дневными грозами имаго либо наблюдались ранним вечером, либо не появлялись вовсе.
Самцы находили места для отдыха в тростнике на высоте 1 м и выше. Когда они замечали добычу, устремлялись к ней, пытаясь поймать. Успешными или неуспешными были их попытки, они всегда возвращались на одну и ту же присаду.

Фото © Владимир Бондарь, Могилевский р-н
Стрекоз, находящихся далеко от места размножения, в основном обнаруживают, когда они отдыхают на деревьях или в кустарниковой растительности; менее 1% особей наблюдали на открытых участках, таких как поля или луга.
Самцы часто обнаруживаются у воды, тогда как самки чаще встречаются далеко от воды (100-900 м), что довольно обычно для стрекоз. Основная биологическая причина этой тенденции, вероятно, заключается в том, что кормление в основном происходит вдали от места размножения, и самкам требуется больше энергии, чем самцам, для гаметогенеза. Кроме того, взрослые самцы максимизируют свою репродуктивную способность, концентрируясь у воды, где возможности для копуляции выше, поскольку прилетевших самок легко заметить и поймать.
Спаривание обычно начинается в полете и завершается сидя на прибрежной растительности; оно длится не менее 10-15 мин, но некоторые авторы указывают не менее 36 мин.
Яйца откладываются в районе произрастания осоки или над открытой поверхностью воды у береговых тростников, реже под нависшими кустами или под высохшими тростниками. Самка откладывает яйца, паря в воздухе, и каждые несколько секунд ударяет о воду кончиком брюшка, высвобождая яйца, часто оставаясь на одном месте в течение нескольких минут. Самец обычно парит поблизости, чтобы отразить вторжение других самцов.
Самцы часто обнаруживаются у воды, тогда как самки чаще встречаются далеко от воды (100-900 м), что довольно обычно для стрекоз. Основная биологическая причина этой тенденции, вероятно, заключается в том, что кормление в основном происходит вдали от места размножения, и самкам требуется больше энергии, чем самцам, для гаметогенеза. Кроме того, взрослые самцы максимизируют свою репродуктивную способность, концентрируясь у воды, где возможности для копуляции выше, поскольку прилетевших самок легко заметить и поймать.
Спаривание обычно начинается в полете и завершается сидя на прибрежной растительности; оно длится не менее 10-15 мин, но некоторые авторы указывают не менее 36 мин.
Яйца откладываются в районе произрастания осоки или над открытой поверхностью воды у береговых тростников, реже под нависшими кустами или под высохшими тростниками. Самка откладывает яйца, паря в воздухе, и каждые несколько секунд ударяет о воду кончиком брюшка, высвобождая яйца, часто оставаясь на одном месте в течение нескольких минут. Самец обычно парит поблизости, чтобы отразить вторжение других самцов.

Фото © Владимир Бондарь, Могилевский р-н
Нимфа. Личинки грязно-желто-коричневатые, грязно-красно-коричневатые или серо-коричневые с темными пятнами, с массивным волосатым телом. Голова довольно маленькая, короткая, приблизительно в 2 раза шире длины, в поперечнике 5 мм, пятиугольная, волосатая; темя выдается довольно сильно, с 3 треугольными пятнами на месте глазков; глаза маленькие, расположены в передних углах головы; от них по направлению к темени идут 2 темные полосы; антенны слабо волосатые, первые два членика утолщенные, 3-й наиболее длинный; затылок большой; внешние стороны его слабо скошены, на каждой стороне среднего шва расположено по 1 четкому почти круглому пятну и более продолговатое пятно вдоль переднего края; задние углы округлые, обрамленные густыми волосками; задний край почти прямой. Маска более яркой окраски, чем остальные части тела, в состоянии покоя достигает середины расстояния между основаниями передней и средней пар ног, широкая, ложковидная, покрывающая все лицевые части. Переднегрудь напоминает воротничок, с выпуклым задним краем; дыхальца среднегруди заметны. Ноги длинные, крепкие, волосатые, с длинными шипами, расположенными рядами; на лапках ряд коротких шипов; коготки довольно длинные и острые, бедра с 2 намечающимися темными кольцами. Брюшко овально-продолговатое, ланцетовидное, постепенно суживающееся к вершине, сильно выпуклое; сегменты почти равные, за исключением Х, очень маленького, образующего род воротничка вокруг анальной пирамиды. Анальная пирамида по длине почти равна IX и Х сегментам, взятым вместе, сильно волосатая; дорсальный придаток немного короче церков, даже почти равен им по длине; церкоиды почти вдвое короче церков.

♀ Фото © Владимир Бондарь, Могилевский р-н
Образ жизни сходен с таковым L. depressa и L. quadrimaculata, но личинки обитают в проточных водах, а не в стоячих (главным образом в медленно текущих, глубоких, заросших камышом ручьях, речках, проточных озерах, прудах), где они живут на дне, зарывшись в илистый или тинистый грунт. Молодые личинки вначале остаются открытыми на дне водоема и только после 3-й линьки прячутся под отмершими частями растений или закапываются в ил и песок. Личинки последних трех возрастов были обнаружены на глубине от 0,7 до 1,5 м.
Из-за своего размера и многолетнего характера водоемы, где развиваются личинки, почти всегда более или менее заселены рыбой. В то время как личинки обычно могут сосуществовать с рыбой без каких-либо проблем, считается, что на них отрицательно влияет наличие роющих донный грунт рыб, например, карпа Cyprinus carpio, и чрезмерные запасы угря. Развитие подводной и плавающей растительности менее важно, чем структурно богатая прибрежная растительность. Кроме того, большинство водоемов должно быть защищено от ветра окружающим лесом.
Личинки вылупляются, по одним данным, через 5-7 недель после откладки яиц, а по другим, уже через 11-12 дней. Так же различаются и данные о числе линек за время личиночного развития, по одним данным, их 11, по другим - 16. Для развития, вероятно, требуется 2 года.
Из-за своего размера и многолетнего характера водоемы, где развиваются личинки, почти всегда более или менее заселены рыбой. В то время как личинки обычно могут сосуществовать с рыбой без каких-либо проблем, считается, что на них отрицательно влияет наличие роющих донный грунт рыб, например, карпа Cyprinus carpio, и чрезмерные запасы угря. Развитие подводной и плавающей растительности менее важно, чем структурно богатая прибрежная растительность. Кроме того, большинство водоемов должно быть защищено от ветра окружающим лесом.
Личинки вылупляются, по одним данным, через 5-7 недель после откладки яиц, а по другим, уже через 11-12 дней. Так же различаются и данные о числе линек за время личиночного развития, по одним данным, их 11, по другим - 16. Для развития, вероятно, требуется 2 года.

♂ Фото © Владимир Бондарь, Могилевский р-н
Отрождение имаго обычно сильно синхронизировано в отдельном водоеме, но может сильно варьироваться в зависимости от температурного баланса соответствующих водоема, отмечены расхождения в сроках в 4-6 недель (в одном и том же году, но в разных водоемах). Через 2-3 недели после начала отрождений в конкретном водоеме активность отрождений резко снижается.
Срок полового созревания молодых особей от 12 до 20 дней.
Средняя продолжительность жизни имаго и сроки их активности различны и зависят от множества факторов, в том числе и от погодных условий данного года. Например, в Чехии на одном и том же водоеме установленная максимальная продолжительность жизни взрослых особей в 2002 г. составила 16 сут., а в 2003 г. - 26 сут. В то же время в 2002 г. активность имаго продолжалась 70 дней, в 2003 г. всего 41 день.
Срок полового созревания молодых особей от 12 до 20 дней.
Средняя продолжительность жизни имаго и сроки их активности различны и зависят от множества факторов, в том числе и от погодных условий данного года. Например, в Чехии на одном и том же водоеме установленная максимальная продолжительность жизни взрослых особей в 2002 г. составила 16 сут., а в 2003 г. - 26 сут. В то же время в 2002 г. активность имаго продолжалась 70 дней, в 2003 г. всего 41 день.

♀ Фото © Владимир Бондарь, Могилевский р-н

Фото © Владимир Бондарь, Могилевский р-н
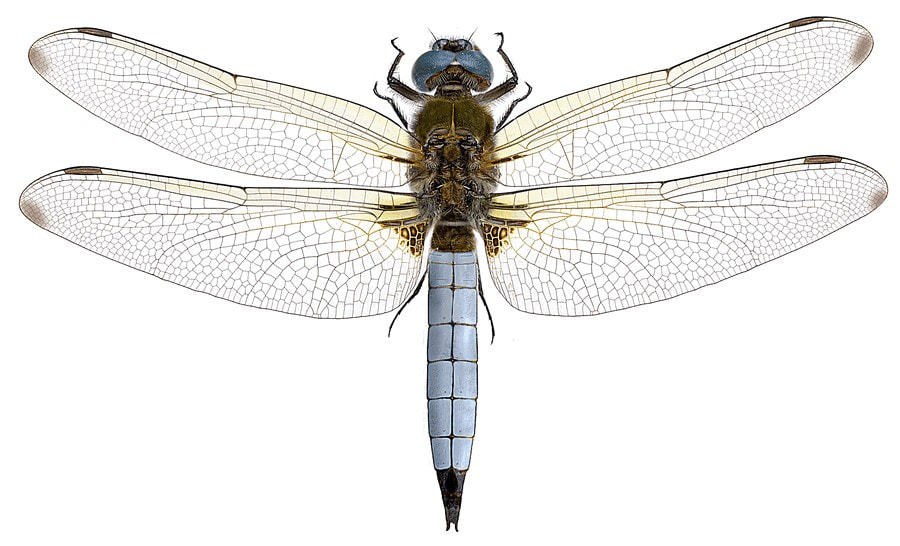
Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0
♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
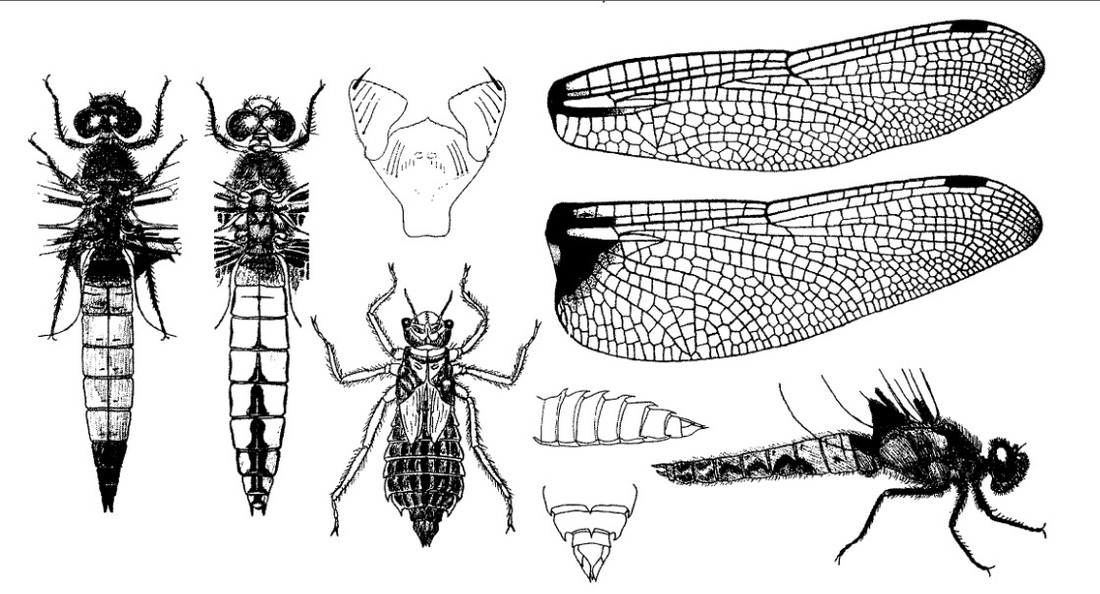
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
4. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Galliani C., Scherini R., Piglia A. "Dragonflies and Damselflies of Europe". WBA Handbooks 7, Verona, 2017: 1-352.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Бондарь В. А. "Libellula fulva Muller, 1764 в Могилевском р-не" / Личное сообщение с фотоматериалом от 26.06.22
9. Smallshire D., Swash A. "Blue Chaser. Libellula fulva" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 256-257.
10. Macagno A. L. M., Boana G., Palestrini C., Stassi M., Rolando A. "Movement and Demographics of Libellula fulva (Odonata, Libellulidae)" / Environ. Entomol. 37(5): 1145-1153. (2008).
11. Dolný A., Matějka P. "A contribution to population biology of Libellula fulva (Odonata: Libellulidae) on coal sludge sedimentation pond (Karviná – Czech Republic)" / Ekológia (Bratislava), Vol. 26, No. 4, p. 341–351, 2007.
12. Schiel F.-J., Mauersberger R. "Libellula fulva Müller, 1764. Spitzenfleck" / Atlas der Libellen Deutschlands (Odonata). Libellula Supplement 14: 278-281. 2015.
13. Mauersberger R., Brauner O. "Spitzenfleck – Libellula fulva Müller 1764" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 122-123.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
4. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Galliani C., Scherini R., Piglia A. "Dragonflies and Damselflies of Europe". WBA Handbooks 7, Verona, 2017: 1-352.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Бондарь В. А. "Libellula fulva Muller, 1764 в Могилевском р-не" / Личное сообщение с фотоматериалом от 26.06.22
9. Smallshire D., Swash A. "Blue Chaser. Libellula fulva" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 256-257.
10. Macagno A. L. M., Boana G., Palestrini C., Stassi M., Rolando A. "Movement and Demographics of Libellula fulva (Odonata, Libellulidae)" / Environ. Entomol. 37(5): 1145-1153. (2008).
11. Dolný A., Matějka P. "A contribution to population biology of Libellula fulva (Odonata: Libellulidae) on coal sludge sedimentation pond (Karviná – Czech Republic)" / Ekológia (Bratislava), Vol. 26, No. 4, p. 341–351, 2007.
12. Schiel F.-J., Mauersberger R. "Libellula fulva Müller, 1764. Spitzenfleck" / Atlas der Libellen Deutschlands (Odonata). Libellula Supplement 14: 278-281. 2015.
13. Mauersberger R., Brauner O. "Spitzenfleck – Libellula fulva Müller 1764" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 122-123.
