СТРЕЛКА ГОЛУБАЯ
Enallagma cyathigerum Charpentier, 1840

♂ Фото © Владимир Бондарь, Могилевский р-н
Enallagma cyathigerum Charpentier, 1840 (стрелка голубая)
Голова широкая, сверху вдвое шире своей длины. На затылке 2 светлых клиновидных пятна. Задний край переднегруди равномерно закругленный или почти плоский, с голубым ободком или голубой точкой по середине. Верхняя часть второго бокового шва груди с черным пятном или с короткой полосой. Бока брюшных тергитов вдоль вентральных швов обычно без черных полос, иногда с очень узкими полосами или с их следами. Не пересекающая плечевой шов светлоокрашенная доплечевая полоса и следующая за ней черная полоса обе хорошо выражены, причем светлая полоса шире расположенной ниже черной полосы. Крылья прозрачные. Птеростигма узкая (равна 1 ячейке), одноцветная, темная. Ноги черные или темно-серые.
♂: Верхняя часть глаз черная, нижняя голубая. Голубой, с черным рисунком. Передние сегменты брюшка почти полностью голубые, с черным сердцевидным пятном в их задней части (которое постепенно увеличивается от III до VII тергита). Темный рисунок на II тергите брюшка состоит только из одного Т-образного пятна (в виде «гриба»), примыкающего к заднему краю членика и доходящего только до середины тергита. Форма рисунка на сегменте II различна, как и степень черного цвета в целом. Только на двух средних сегментах черные пятна продолговатые и занимают около половины длины сегментов. Конец брюшка (тергиты VIII-IX) целиком голубой, кроме двух маленьких черных пятен на IX и за исключением придатков. Голубой цвет становится серым при низкой температуре. Нижние анальные придатки длиннее верхних, узкотреугольные.
♀: Верхняя часть глаз черная, нижняя зависит от цветовой формы. Имеет две цветовые формы - голубой и тускло-зеленой окраски, с возрастом становящейся светло-коричневой. В отличие от самцов у самок продолговатое черное пятно развито на каждом брюшном сегменте. Формы черных отметин на брюшке включают «чертополох» на сегменте II (проходит от переднего до заднего края сегмента), различные формы «ракеты» на сегментах III-VII и треугольник на сегменте VIII. На VIII стерните брюшка перед яйцекладом имеется шип.
У ювенильных особей светлые участки изначально имеют соломенный цвет.
♂: Верхняя часть глаз черная, нижняя голубая. Голубой, с черным рисунком. Передние сегменты брюшка почти полностью голубые, с черным сердцевидным пятном в их задней части (которое постепенно увеличивается от III до VII тергита). Темный рисунок на II тергите брюшка состоит только из одного Т-образного пятна (в виде «гриба»), примыкающего к заднему краю членика и доходящего только до середины тергита. Форма рисунка на сегменте II различна, как и степень черного цвета в целом. Только на двух средних сегментах черные пятна продолговатые и занимают около половины длины сегментов. Конец брюшка (тергиты VIII-IX) целиком голубой, кроме двух маленьких черных пятен на IX и за исключением придатков. Голубой цвет становится серым при низкой температуре. Нижние анальные придатки длиннее верхних, узкотреугольные.
♀: Верхняя часть глаз черная, нижняя зависит от цветовой формы. Имеет две цветовые формы - голубой и тускло-зеленой окраски, с возрастом становящейся светло-коричневой. В отличие от самцов у самок продолговатое черное пятно развито на каждом брюшном сегменте. Формы черных отметин на брюшке включают «чертополох» на сегменте II (проходит от переднего до заднего края сегмента), различные формы «ракеты» на сегментах III-VII и треугольник на сегменте VIII. На VIII стерните брюшка перед яйцекладом имеется шип.
У ювенильных особей светлые участки изначально имеют соломенный цвет.

♀ Фото © Виталий Сирота, г. Минск
Считается самой голубой из равнокрылых стрекоз. Оба пола легко отличить от других голубых стрелок по широким предплечевым полосам и отсутствию «шпоры» сбоку на груди. У самцов голубые предплечевые полосы намного шире, чем у видов Coenagrion. Бока груди почти полностью голубые, а черная отметка на II сегменте брюшка редко выходит за пределы срединного округлого пятна, обычно соединенного с задним межсегментарным кольцом короткой ножкой. Самка встречается в двух цветовых формах: одна черная с зеленоватыми переднеплечевыми полосами и зеленоватыми боками груди и брюшка, а другая черная с ярко-голубыми отметинами, хотя на брюшке они менее обширны, чем у самца. У обеих форм генитальный шип присутствует на вентральной поверхности VIII сегмента брюшка, в отличие от самок Coenagrion.
Длина 29-36 мм, брюшко 22-28 мм, заднее крыло 18-21 мм.
Лет: май - сентябрь (октябрь). Основной период отрождения имаго - с середины мая до начала июля, а основное время лёта приходится на конец мая - середину августа. В отдельные годы имеются поздние наблюдения молодых имаго с конца августа, которые указывают на развитие второго поколения в эти годы. Вероятно, именно этих имаго иногда можно увидеть в небольшом количестве до конца сентября или начала октября.
Длина 29-36 мм, брюшко 22-28 мм, заднее крыло 18-21 мм.
Лет: май - сентябрь (октябрь). Основной период отрождения имаго - с середины мая до начала июля, а основное время лёта приходится на конец мая - середину августа. В отдельные годы имеются поздние наблюдения молодых имаго с конца августа, которые указывают на развитие второго поколения в эти годы. Вероятно, именно этих имаго иногда можно увидеть в небольшом количестве до конца сентября или начала октября.

© Семён Левый, Кобринский р-н, Брестская обл.
Как правило, в естественных популяциях сосуществуют два фенотипа взрослых самок: одна морфа окрашена как конспецифический самец (андроморфная), а другая морфа отличается окраской тела (гетероморфная). Большинство современных объяснений, касающихся сохранения этого полиморфизма, указывают на сексуальный конфликт как на главную силу отбора. В то время как самцы стремятся максимизировать частоту совокупления, самки развили черты, позволяющие избегать чрезмерного домогательства. Одной из таких адаптивных черт может быть окраска самок, при котором определенные фенотипы выигрывают, подвергаясь меньшему преследованию со стороны ищущих себе пару самцов, которые не сразу узнают в них самку.
Временное изменение наблюдаемых пропорций для вида в Бельгии составляло от 0% до 40% самок, являющихся андроморфными. Однако в разных странах Европы отмечены различия в доле андроморфных самок этого вида в диапазоне от 17 до 76%. Пропорции морф самок различаются не только в зависимости от популяции или года, но и в более коротких временных масштабах для одного водоема. Для изучаемых в Бельгии популяций пропорции андроморфных самок колебались на суточном уровне, но оставались стабильными в течение всего сезона. Тем не менее, сезонные колебания могут иметь место в других популяциях.
Временное изменение наблюдаемых пропорций для вида в Бельгии составляло от 0% до 40% самок, являющихся андроморфными. Однако в разных странах Европы отмечены различия в доле андроморфных самок этого вида в диапазоне от 17 до 76%. Пропорции морф самок различаются не только в зависимости от популяции или года, но и в более коротких временных масштабах для одного водоема. Для изучаемых в Бельгии популяций пропорции андроморфных самок колебались на суточном уровне, но оставались стабильными в течение всего сезона. Тем не менее, сезонные колебания могут иметь место в других популяциях.

♂ Фото © Виталий Сирота, г. Минск
Заселяет относительно крупные стоячие водоемы: озера, пруды, речные старицы с чистой водой, чаще, но не обязательно, с богатой водной растительностью и поясом прибрежных тростников. Предпочитают большие пруды и озера или медленно текущие реки. В отличие от C. puella, редко размножается в очень маленьких прудах.
Спектр местообитаний очень широк. Предпочтение отдается средним и крупным мезо- и эвтрофным водоемам с открытой водной поверхностью. Для равнины Верхнего Рейна как необходимое условие для обитания вида указывается свободная водная поверхность не менее 80 м². В центре внимания вида находятся постоянные стоячие водоемы различного происхождения: естественные озера, затопленные карьеры, водохранилища, пруды. Также заселяются богатые питательными веществами небольшие водоемы, болотные пруды и торфяники. Кроме того, вид также встречается в медленно текущих канавах и каналах или заводях со спокойным течением проточных водотоков.
В Бранденбурге (Германия) считается типичным видом для озер, откуда поступает более половины всех регистраций. Следующими указываются пруды и грунтовые воды в водоемах всех видов, в то время как водотоки играют очень второстепенную роль в качестве среды обитания. Подчеркивается предпочтение больших открытых акваторий. Один из видов, способных оптимально заселять водоемы, богатые рыбой.
В то же время, для Британии и Ирландии указывается избегание маленьких прудов и канав, а также водоемов, населенных рыбами. В Испании указывается размножение не только в маленьких, но и во временных прудах, а также солоноватых стоячих водоемах, если они хорошо освещаются солнцем.
Спектр местообитаний очень широк. Предпочтение отдается средним и крупным мезо- и эвтрофным водоемам с открытой водной поверхностью. Для равнины Верхнего Рейна как необходимое условие для обитания вида указывается свободная водная поверхность не менее 80 м². В центре внимания вида находятся постоянные стоячие водоемы различного происхождения: естественные озера, затопленные карьеры, водохранилища, пруды. Также заселяются богатые питательными веществами небольшие водоемы, болотные пруды и торфяники. Кроме того, вид также встречается в медленно текущих канавах и каналах или заводях со спокойным течением проточных водотоков.
В Бранденбурге (Германия) считается типичным видом для озер, откуда поступает более половины всех регистраций. Следующими указываются пруды и грунтовые воды в водоемах всех видов, в то время как водотоки играют очень второстепенную роль в качестве среды обитания. Подчеркивается предпочтение больших открытых акваторий. Один из видов, способных оптимально заселять водоемы, богатые рыбой.
В то же время, для Британии и Ирландии указывается избегание маленьких прудов и канав, а также водоемов, населенных рыбами. В Испании указывается размножение не только в маленьких, но и во временных прудах, а также солоноватых стоячих водоемах, если они хорошо освещаются солнцем.

♂♂ Фото © Виктор Рындевич, Смолевичский р-н, Минская обл.
Что касается ежедневной фенологии, то вид можно обнаружить у воды в течение всего дня с 8:30 до 20:00, если позволяет погода, с максимальной активностью во второй половине дня. Самцы, готовые к размножению, обычно сидят у края открытой водной поверхности, на торчащих из воды частях подводных растений, старом тростнике или обломках растений. Оттуда они совершают относительно длительные патрульные полеты низко над поверхностью воды.
В солнечную погоду может встречаться в очень больших количествах и роиться низко над поверхностью воды, часто далеко от краев водоема, периодически садясь на надводные головки цветков водных растений, сомкнув крылья над брюшком или слегка приоткрыв их. Неполовозрелые особи часто разлетаются далеко от водоемов.
Встречается совместно со многими другими видами стрекоз, которые имеют схожие места обитания. Ночует на более широких стеблях, чем Ischnura elegans или C. puella. В прохладную или ветреную погоду могут быть потревожены целые полчища стрекоз, укрывающихся в относительного спокойной высокой маргинальной растительности.
Половозрелая взрослая жизнь имаго составляет около 12 дней, но самцы иногда могут жить до 39 дней.
В солнечную погоду может встречаться в очень больших количествах и роиться низко над поверхностью воды, часто далеко от краев водоема, периодически садясь на надводные головки цветков водных растений, сомкнув крылья над брюшком или слегка приоткрыв их. Неполовозрелые особи часто разлетаются далеко от водоемов.
Встречается совместно со многими другими видами стрекоз, которые имеют схожие места обитания. Ночует на более широких стеблях, чем Ischnura elegans или C. puella. В прохладную или ветреную погоду могут быть потревожены целые полчища стрекоз, укрывающихся в относительного спокойной высокой маргинальной растительности.
Половозрелая взрослая жизнь имаго составляет около 12 дней, но самцы иногда могут жить до 39 дней.

♂ Фото © Владимир Бондарь, Быховский р-н, Могилевская обл.
Описывается нетипичный и, можно сказать, уникальный случай из Баварии, когда наблюдалась активная подводная охота самки (Weihrauch, 2002). Нетипичным этот случай является потому, что самка стрекозы охотилась на больших ивовых тлей Tuberolachnus salignus, которые живут на ветках ивы и вовсе не под водой. Свежая ветка с зелеными листьями была зажата под водой намокшей толстой доской и была плотно усеяна тлями. Осталось невыясненным, обнаружила добычу стрекоза случайно, пытаясь сделать на ветке яйцекладку, либо охотилась специально. Самка села на ветку и начала спускаться по ней под воду головой вперед. Стрекоза, чья голова и грудь тем временем были полностью погружены в воду, начала поедать обессилевших и полуутонувших тлей, откусывая, а затем съедая. Она методично ощипывала ветку и за 5 мин. переместилась вниз на 4-5 см, съев 7-8 тлей. Так как ветка лежала под водой под углом около 30°, несмотря на перемещение, стрекоза всегда держала заднюю часть брюшка над водой. В конце концов она переключилась на нижнюю сторону ветки, изогнув брюшко так, что последние два сегмента торчали над поверхностью воды, как трубка. В этот момент наблюдатель попытался немного повернуть ветку для лучшего обзора, но неудачно. Ветка выскользнула из-под доски и катапультировала стрекозу по широкой дуге из поля зрения.

♀ Фото © Семён Левый, Малоритский р-н, Брестская обл.
Самцы крепкие и агрессивные. Патрулируют водоем, летая низко над поверхностью воды, гоняясь за самками, а также в ожидании самок, выходящих из воды после откладки яиц, которых «охраняют» и помогают им вернуться на берег. Отмечается межвидовая конкуренция между E. cyathigerum и Erythromma najas.
Копуляция длится 20 минут и более и часто происходит вдали от воды.
Яйца откладываются «в тандеме» в поверхностную растительность, цветоножки и листья Potamogeton natans и в цветоножки Myriophyllum. Впрочем, для некоторых мест наличие именно этих растений некритично, нередко указывается лишь наличие участков с водной растительностью, хотя бы выборочно достигающей поверхности воды. Эта растительность необходима для посадки самок, по которой они могут опускаться на глубину для откладки яиц, а также для личинок, которые покидают воду для отрождения в имаго.
Самка при этом процессе погружает свое брюшко в воду, а иногда погружается и целиком и остается под водой довольно продолжительное время (до 90 минут). При опускании самки в воду самец обычно отпускает ее и улетает. Хотя согласно другим авторам, самец отсоединяется и ждет ее появления, охраняя точку спуска самки под воду, летая, либо дежуря рядом, усевшись на какой-либо небольшой стебель растения над поверхностью воды. Вероятно, степень «охраны» самцом своей партнерши напрямую зависит от плотности популяции на данном участке.
При высокой плотности конкуренция между самцами очень острая. Летающие самцы с готовностью преследуют любые пролетающие объекты, которые «выглядят» как самки, например семена с хохолком («парашютиком») цветков сложноцветных. Тандемные пары, приближающиеся к основным местам откладки яиц, сразу же подвергаются преследованию со стороны одиночных самцов.
Копуляция длится 20 минут и более и часто происходит вдали от воды.
Яйца откладываются «в тандеме» в поверхностную растительность, цветоножки и листья Potamogeton natans и в цветоножки Myriophyllum. Впрочем, для некоторых мест наличие именно этих растений некритично, нередко указывается лишь наличие участков с водной растительностью, хотя бы выборочно достигающей поверхности воды. Эта растительность необходима для посадки самок, по которой они могут опускаться на глубину для откладки яиц, а также для личинок, которые покидают воду для отрождения в имаго.
Самка при этом процессе погружает свое брюшко в воду, а иногда погружается и целиком и остается под водой довольно продолжительное время (до 90 минут). При опускании самки в воду самец обычно отпускает ее и улетает. Хотя согласно другим авторам, самец отсоединяется и ждет ее появления, охраняя точку спуска самки под воду, летая, либо дежуря рядом, усевшись на какой-либо небольшой стебель растения над поверхностью воды. Вероятно, степень «охраны» самцом своей партнерши напрямую зависит от плотности популяции на данном участке.
При высокой плотности конкуренция между самцами очень острая. Летающие самцы с готовностью преследуют любые пролетающие объекты, которые «выглядят» как самки, например семена с хохолком («парашютиком») цветков сложноцветных. Тандемные пары, приближающиеся к основным местам откладки яиц, сразу же подвергаются преследованию со стороны одиночных самцов.

© Семён Левый, Жабинковский р-н, Брестская обл.
Самки в тандеме являются пассивной стороной при выборе места. «Руководят» тандемом самцы, пытаясь избежать внимания других самцов. Когда пара приближается к воде, самка с готовностью схватывает любое растение, которое торчит над поверхностью воды. Одиночные самцы неоднократно беспокоят тандемную пару, пытаясь схватить самку ногами. Чаще всего это происходит, когда самка находится на поверхности воды или близко к ней. Однако имеется несколько наблюдений попыток одиночных самцов сделать это, пока тандем все еще находился в полете.
При высокой плотности самцы заинтересованы оставаться в тандеме с самкой как можно дольше. Это увеличивает шансы охранять самку, пока она не отложит яйца, которые, вероятно, оплодотворил данный самец. Это поведение охраны необходимо при высокой плотности, поскольку самцы-агрессоры, похоже, идут на многое, чтобы «завоевать» самку, и готовы погружаться под воду, чтобы достичь тандема.
Например, в одной паре самец оставался в тандеме, хотя самка начала откладывать яйца уже под воду. Он оставался удерживать партнершу в течение 43 сек, прежде чем отпустить ее, чтобы та продолжила яйцекладку самостоятельно. У другой тандемной пары, при контакте с водой самка сразу же погрузилась в воду, а самец все еще находился в тандеме. Он отпустил самку в тот момент, когда верхняя часть его груди погрузилась в воду. В этот момент прилетел одиночный самец и попытался схватить ногами почти погруженную в воду самку. Первый самец повторил попытку соединиться в тандем с самкой, но последовала борьба, в которой агрессор смог оттеснить первого самца от самки. В этот момент самка была под водой и удерживалась обоими самцами. Первый самец всплыл и улетел. После чего второй самец начал опускаться по задней части брюшка самки, которая была полностью под водой и пыталась отложить яйца. Достигнув соединения в тандем, самец рывками смог оттащить самку от стебля растения и подняться вместе с ней на поверхность воды. Первоначально тандем не мог взлететь из-за переувлажнения тела и крыльев, но самцу удалось подтащить самку к ближайшему надводному стеблю. После нескольких попыток взлететь, это им удалось и они улетели тандемом.
При высокой плотности самцы заинтересованы оставаться в тандеме с самкой как можно дольше. Это увеличивает шансы охранять самку, пока она не отложит яйца, которые, вероятно, оплодотворил данный самец. Это поведение охраны необходимо при высокой плотности, поскольку самцы-агрессоры, похоже, идут на многое, чтобы «завоевать» самку, и готовы погружаться под воду, чтобы достичь тандема.
Например, в одной паре самец оставался в тандеме, хотя самка начала откладывать яйца уже под воду. Он оставался удерживать партнершу в течение 43 сек, прежде чем отпустить ее, чтобы та продолжила яйцекладку самостоятельно. У другой тандемной пары, при контакте с водой самка сразу же погрузилась в воду, а самец все еще находился в тандеме. Он отпустил самку в тот момент, когда верхняя часть его груди погрузилась в воду. В этот момент прилетел одиночный самец и попытался схватить ногами почти погруженную в воду самку. Первый самец повторил попытку соединиться в тандем с самкой, но последовала борьба, в которой агрессор смог оттеснить первого самца от самки. В этот момент самка была под водой и удерживалась обоими самцами. Первый самец всплыл и улетел. После чего второй самец начал опускаться по задней части брюшка самки, которая была полностью под водой и пыталась отложить яйца. Достигнув соединения в тандем, самец рывками смог оттащить самку от стебля растения и подняться вместе с ней на поверхность воды. Первоначально тандем не мог взлететь из-за переувлажнения тела и крыльев, но самцу удалось подтащить самку к ближайшему надводному стеблю. После нескольких попыток взлететь, это им удалось и они улетели тандемом.

© Владимир Бондарь, Быховский р-н, Могилевская обл.
Нимфа. Окраска тела желто-коричневая, но чаще всего светло-зеленая, с неясно выраженным рисунком. Тело гладкое, длиной 19-20 мм и шириной 3-4 мм. Голова широкая, 4 мм в поперечнике, но относительно короткая в продольном направлении от лба до затылка, с темными и светлыми отметинами на вершине; глаза эллипсоидной формы, темные, с мало выдающимися внешними краями; антенны с темными кольцами; затылок относительно большой, с сильно скошенными внешними краями, округлыми задними углами, снабженными довольно большими хорошо заметными шипиками и с очень вогнутым задним краем. Маска в состоянии покоя доходит до основания передней пары ног; подбородок очень узкий у основания, но сильно расширенный у вершины. Переднегрудь короткая, с относительно острыми и темными углами. Крыловые чехлы прямые, длиной около 5 мм, доходят до верхней половины IV сегмента. Ноги довольно длинные, передние 7 мм, средние 8 мм, задние 11 мм; на бедрах ближе к дистальному краю по темному кольцу, не всегда заметному; ноги вооружены острыми длинными и короткими шипами, расположенными продольными рядами. Каждый сегмент брюшка, за исключением Х сегмента, с темным подковообразным пятном и со светлым пятном внутри подковы; светлые пятна рассечены узкой еще более светлой полоской; внешние края светлее; межсегментальные пространства с 2 очень темными пятнами. Хвостовые жаберные пластинки желтовато-коричневые, эллипсоидной формы, островершанные или округловершинные, широкие, 1,5-1,7 мм, длиной 5,5-6 мм, сужение к острию начинается значительно дистальнее середины; темные поперечные полосы пересекают пластинки; трахей много, и разветвление хорошо видимо. Створки яйцеклада самки островершинные, простираются почти до дистального края Х стернита; субанальные пластинки самца и самки резко отличаются друг от друга. У самки они треугольные, островершинные; у самца - округлые.

♂ Фото © Виталий Сирота, г. Бобруйск, Могилевская обл.
Обитают главным образом в крупных стоячих водоемах (большие озера, пруды, канавы), но встречаются и в заводях медленно текущих рек и даже в солоноватых водах. Типичные фитофилы, живут в зарослях погруженных макрофитов на глубине 0,2-1 м. Необходимыми структурами среды обитания являются подводная водная растительность, доходящая до поверхности воды, и хотя бы фрагментарные заросли берегового тростника для вертикальных береговых присад. Присутствие рыбы не препятствуют воспроизводству популяций вида, если имеется достаточное количество подводной растительности, в которой могут укрыться личинки. Например, в 28 водоемах с E. cyathigerum, исследованных в Верхнем Рейне, почти во всех были обнаружены рыбные запасы. По отношению к гидрохимическим факторам, таким как значение рН, электропроводность и жесткость воды, вид оказывается индифферентным в очень широком диапазоне. 50% личинок популяции переживают резкое падение содержания кислорода в пруду в течение нескольких дней. Также продемонстрировано успешное размножение в солоноватой воде.
Личинки развиваются от 1 до 3 лет, обычно 2 года. Однако сроки развития отличаются не только географически, но и в зависят от конкретного лета в данном году. Например, в Германии вид считается моновольтинным (развитие за 1 год), но в годы с благоприятными погодными условиями возможно бивольтинное развитие, т. е. отрождение в двух поколениях за год. В отдельные годы имеются поздние наблюдения молодых имаго с конца августа, которые указывают на развитие второго поколения в эти годы. Вероятно, именно этих имаго иногда можно увидеть в небольшом количестве до конца сентября или начала октября.
Личинки развиваются от 1 до 3 лет, обычно 2 года. Однако сроки развития отличаются не только географически, но и в зависят от конкретного лета в данном году. Например, в Германии вид считается моновольтинным (развитие за 1 год), но в годы с благоприятными погодными условиями возможно бивольтинное развитие, т. е. отрождение в двух поколениях за год. В отдельные годы имеются поздние наблюдения молодых имаго с конца августа, которые указывают на развитие второго поколения в эти годы. Вероятно, именно этих имаго иногда можно увидеть в небольшом количестве до конца сентября или начала октября.

♂ Фото © Семён Левый, Кобринский р-н, Брестская обл.

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
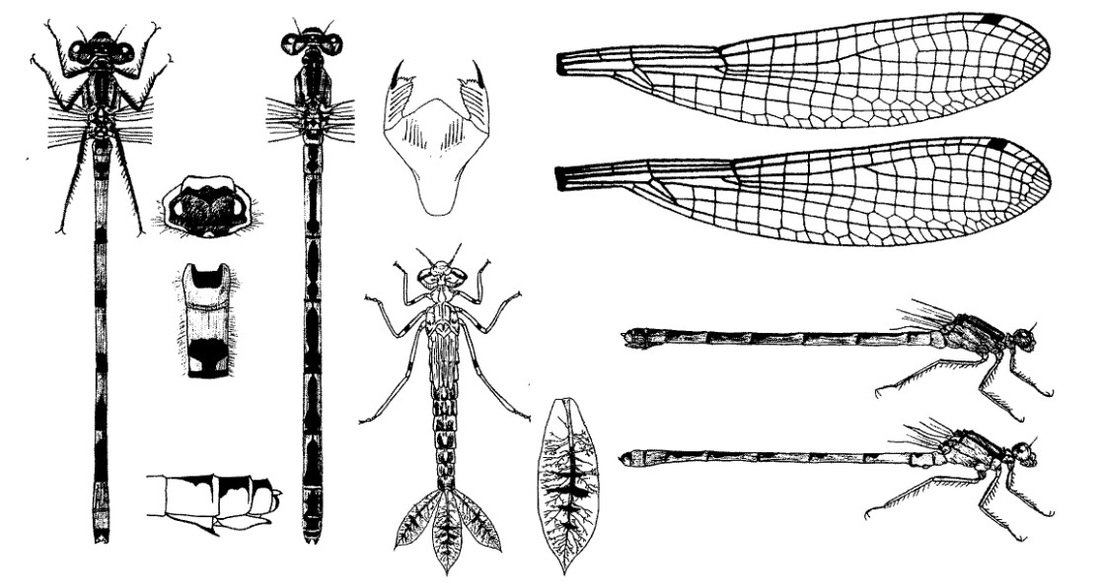
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Common Bluet. Enallagma cyathigerum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 68-69.
9. Moreno-Benítez J. M., Ripoll J. "Enallagma cyathigerum" / in: Dragonflies along the Great Málaga Path and in the Province. Identification Guide, 2018. pp. 74-75.
10. Smallshire D., Swash A. "Common Blue Damselfly. Enallagma cyathigerum" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 90-91.
11. Bots J., De Bruyn L., Adriaens T., Dumont H., Robby Stoks R., Van Gossum H. "Seasonal and diurnal variation in the proportions of female morphs of the damselfly Enallagma cyathigerum" / Animal Biology, Volume 57, Issue 2, 2007: 217-230.
12. Haacks M., Brockhaus T., Mauersberger R., Willigalla C. "Enallagma cyathigerum (Charpentier, 1840). Gemeine Becherjungfer, Becher-Azurjungfer" / Libellula Supplement 14: 94-97.
13. Weihrauch F. "Ein Weibchen von Enallagma cyathigerum als Unterwasser-Prädator der Großen Weidenrindenlaus (Odonata: Coenagrionidae; Homoptera: Lachnidae)" / Libellula 21 (3/4 ): 175-180. 2002.
14. Cham S. "Mate guarding behaviour during intense competition for females in the Common Blue Damselfiy Enallagma cyathigerum (Charpentier)" / Journal of the British Dragonfly Society, Vol. 18, No. 1-2, 2002. pp. 46-48.
15. Mauersberger R., Willigalla C. "Becher-Azurjungfer – Enallagma cyathigerum (Charpentier 1840)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 48-49.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Common Bluet. Enallagma cyathigerum" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 68-69.
9. Moreno-Benítez J. M., Ripoll J. "Enallagma cyathigerum" / in: Dragonflies along the Great Málaga Path and in the Province. Identification Guide, 2018. pp. 74-75.
10. Smallshire D., Swash A. "Common Blue Damselfly. Enallagma cyathigerum" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 90-91.
11. Bots J., De Bruyn L., Adriaens T., Dumont H., Robby Stoks R., Van Gossum H. "Seasonal and diurnal variation in the proportions of female morphs of the damselfly Enallagma cyathigerum" / Animal Biology, Volume 57, Issue 2, 2007: 217-230.
12. Haacks M., Brockhaus T., Mauersberger R., Willigalla C. "Enallagma cyathigerum (Charpentier, 1840). Gemeine Becherjungfer, Becher-Azurjungfer" / Libellula Supplement 14: 94-97.
13. Weihrauch F. "Ein Weibchen von Enallagma cyathigerum als Unterwasser-Prädator der Großen Weidenrindenlaus (Odonata: Coenagrionidae; Homoptera: Lachnidae)" / Libellula 21 (3/4 ): 175-180. 2002.
14. Cham S. "Mate guarding behaviour during intense competition for females in the Common Blue Damselfiy Enallagma cyathigerum (Charpentier)" / Journal of the British Dragonfly Society, Vol. 18, No. 1-2, 2002. pp. 46-48.
15. Mauersberger R., Willigalla C. "Becher-Azurjungfer – Enallagma cyathigerum (Charpentier 1840)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 48-49.
