СТРЕЛКА-ДЕВУШКА
Coenagrion puella Linnaeus, 1758

♂ Фото © Владимир Бондарь, Могилевский р-н
Coenagrion puella Linnaeus, 1758 (стрелка-девушка)
Голова широкая, сверху вдвое шире своей длины, темная сверху. Между крупными затылочными пятнами светлой полосы нет; иногда имеются две точки или начало линии. Задний край переднеспинки с тремя примерно одинаково развитыми лопастями. Лопасти обособлены слабо. Верх груди темный, на боку светлой груди короткая черная линия, светлые предплечевые полосы уже черной линии внизу. Кроме того, имеется дополнительная черная линия («шпора»), идущая от основания переднего крыла на одну треть расстояния по боковой стороны груди к ногам. Крылья прозрачные. Птеростигма узкая (равна 1 ячейке). Птеростигма у обоих полов одноцветная, черная, серая или бледная. Ноги черные или темно-серые.
♂: Основной цвет голубой. Черные пятна на голубом брюшке распределенными следующим образом: базальное черное кольцо на II-VII сегментах, на сегментах VI и VII неполное; на дорсальной стороне сегмента II черное U-образное пятно, которое обычно не соединяется с основанием указанного сегмента, но это пятно сильно варьирует по толщине; сегменты III-V имеют дорсально черные пятна у основания, от которых вдоль сегмента сбоку отходят две тонкие линии; в некоторых случаях эти линии могут не быть связаны с базальной меткой сегмента III; на VI сегменте базальное пятно занимает почти половину, с небольшим зубцом в его средней задней части и толстыми боковыми линиями; сегмент VII сверху черный, а сегмент VIII полностью голубой, хотя у некоторых особей имеются черные точки или линии; IX сегмент голубой, с очень изменчивым черным рисунком в его основании (чаще парные или сросшиеся черные отметки в виде галстука-бабочки), Х сегмент черный на большей части, но снизу голубой. Верхние анальные придатки расходятся в основании, с направленным внутрь вершинным зубцом; дорсальный выступ нижних придатков мощный.
♀: Основной цвет зеленоватый, реже голубовато-серый, с широкими черными пятнами на дорсальной поверхности брюшка, так что сверху каждый сегмент выглядит черным с зеленоватой окантовкой. Тергиты III-X черные, III-VIII (или IV-VII) с узким голубым пояском у переднего края; II тергит с черным пятном через весь членик. Самки также встречаются в голубой форме, у которой голубой цвет простирается на менее 30% на сегментах IV-V.
У неполовозрелых особей голубые участки заменены на беловатые цвета.
♂: Основной цвет голубой. Черные пятна на голубом брюшке распределенными следующим образом: базальное черное кольцо на II-VII сегментах, на сегментах VI и VII неполное; на дорсальной стороне сегмента II черное U-образное пятно, которое обычно не соединяется с основанием указанного сегмента, но это пятно сильно варьирует по толщине; сегменты III-V имеют дорсально черные пятна у основания, от которых вдоль сегмента сбоку отходят две тонкие линии; в некоторых случаях эти линии могут не быть связаны с базальной меткой сегмента III; на VI сегменте базальное пятно занимает почти половину, с небольшим зубцом в его средней задней части и толстыми боковыми линиями; сегмент VII сверху черный, а сегмент VIII полностью голубой, хотя у некоторых особей имеются черные точки или линии; IX сегмент голубой, с очень изменчивым черным рисунком в его основании (чаще парные или сросшиеся черные отметки в виде галстука-бабочки), Х сегмент черный на большей части, но снизу голубой. Верхние анальные придатки расходятся в основании, с направленным внутрь вершинным зубцом; дорсальный выступ нижних придатков мощный.
♀: Основной цвет зеленоватый, реже голубовато-серый, с широкими черными пятнами на дорсальной поверхности брюшка, так что сверху каждый сегмент выглядит черным с зеленоватой окантовкой. Тергиты III-X черные, III-VIII (или IV-VII) с узким голубым пояском у переднего края; II тергит с черным пятном через весь членик. Самки также встречаются в голубой форме, у которой голубой цвет простирается на менее 30% на сегментах IV-V.
У неполовозрелых особей голубые участки заменены на беловатые цвета.

© Виталий Сирота, г. Минск
У самцов голубые предплечевые полосы на груди более узкие, чем у Enallagma cyathigerum и других обычных голубых равнокрылых стрекоз, а черные отметки на II сегменте брюшка совершенно другие, обычно представляют собой тонкую U-образную форму. Подобно другим видам Coenagrion, имеет две короткие толстые черные линии на боках груди у обоих полов, в то время как у аналогичного вида E. cyathigerum такая линия только одна. Самки могут встречаться в одной из нескольких форм: одна темная с зеленоватыми отметинами на груди и брюшке, а другие формы имеют голубые отметины, которые могут быть довольно обширными и очень напоминать самок C. pulchellum, от которых их можно отличить по форме заднего края переднеспинки, которая не трехлопастная, как у C. pulchellum. Кроме того, голубой цвет на сегментах IV-V простирается менее чем на 30%, а у самок C. pulchellum - более 30%.
Длина 33-35 мм, брюшко 22-31 мм, заднее крыло 16-22 мм.
Лет: конец мая - начало сентября, но чаще всего встречается в начале лета.
Длина 33-35 мм, брюшко 22-31 мм, заднее крыло 16-22 мм.
Лет: конец мая - начало сентября, но чаще всего встречается в начале лета.

♂ Фото © Виталий Сирота, Минский р-н
Обитает повсеместно вблизи рек, озер, ручьев, пойменных озер и прудов, предпочитая более мелкие и защищенные участки. Развивается во всех типах стоячих водоемов, но преимущественно в более мелких, таких как пруды и болота. Обязательным условием является четко выраженная зона плавающих лиственных растений и структурно богатая прибрежная растительность. Часто заселяются также болота, болотные канавы и торфяные разрезы. Реже заселяет или отсутствует вовсе в карьерах с небольшой прибрежной растительностью или озерах с крутыми берегами. Медленно текущие водотоки, такие как крупные реки и луговые канавы, также регулярно заселяются видом при соответствующем развитии растительности.
Вид с более или менее динамичной численностью. В отдельные годы вследствие заморов численность сокращается, затем снова восстанавливается. Может встречаться совместно почти со всеми другими видами, которые разделяют его географический ареал.
Пик численности взрослых особей достигается примерно через 2 недели после массового отрождения, когда половозрелые особи возвращаются к водоему для размножения. Показатели обнаружений значительно падают в начале августа, а последние имаго часто наблюдаются в середине августа. Обнаружения в сентябре, часто после недельной паузы, являются исключениями.
Обычно стрекозы этого вида сидят на околоводной и водной растительности. Летают медленно, лишь на короткие расстояния, редко улетает далеко на большие участки открытой воды (в отличие от Enallagma cyathigerum). Особи данного вида боятся ветра, поэтому летают среди растительности. Взрослые особи сидят на более широких стеблях, чем Ischnura elegans, но менее широких, чем E. cyathigerum, и только их глаза видны с обеих сторон спереди. Это защитный механизм: они могут передвигаться вокруг стебля при приближении опасности.
Вид с более или менее динамичной численностью. В отдельные годы вследствие заморов численность сокращается, затем снова восстанавливается. Может встречаться совместно почти со всеми другими видами, которые разделяют его географический ареал.
Пик численности взрослых особей достигается примерно через 2 недели после массового отрождения, когда половозрелые особи возвращаются к водоему для размножения. Показатели обнаружений значительно падают в начале августа, а последние имаго часто наблюдаются в середине августа. Обнаружения в сентябре, часто после недельной паузы, являются исключениями.
Обычно стрекозы этого вида сидят на околоводной и водной растительности. Летают медленно, лишь на короткие расстояния, редко улетает далеко на большие участки открытой воды (в отличие от Enallagma cyathigerum). Особи данного вида боятся ветра, поэтому летают среди растительности. Взрослые особи сидят на более широких стеблях, чем Ischnura elegans, но менее широких, чем E. cyathigerum, и только их глаза видны с обеих сторон спереди. Это защитный механизм: они могут передвигаться вокруг стебля при приближении опасности.

♀ (андроморфная) Фото © Владимир Бондарь, Могилевский р-н
Не является территориальным видом, но есть свидетельства того, что его плотность в некоторой степени регулируется поведением, которое включает демонстрацию угрозы. Спаривание продолжительное и происходит только в теплые солнечные дни. Успех спаривания самок зависит от продолжительности их жизни: скорость яйцекладки и размер кладки имеют меньшее значение для вида.
Яйцекладка происходит в тандеме с вертикально расположенным самцом, иногда самка полностью погружается в воду. Яйца откладываются самкой на нижнюю сторону плавающих и погруженных в воду листьев водных растений: рдестов, кубышек и др., а также в ткани мертвых растений и затонувших стволов и веток деревьев. Яйца располагаются прямыми или концентрическими рядами.
Интересно, что при исследовании размера кладки и морфометрии яиц трех цветовых морф самок (андроморфных - голубых, гетероморфных - зеленых и промежуточных - зеленая грудь, голубое брюшко), как то и ожидалось, кладки не различались ни по количеству, ни по размеру яиц. Однако обнаружено, что промежуточная морфа имеет более удлиненные яйца по сравнению с гетеро- и андроморфными самками.
Яйцекладка происходит в тандеме с вертикально расположенным самцом, иногда самка полностью погружается в воду. Яйца откладываются самкой на нижнюю сторону плавающих и погруженных в воду листьев водных растений: рдестов, кубышек и др., а также в ткани мертвых растений и затонувших стволов и веток деревьев. Яйца располагаются прямыми или концентрическими рядами.
Интересно, что при исследовании размера кладки и морфометрии яиц трех цветовых морф самок (андроморфных - голубых, гетероморфных - зеленых и промежуточных - зеленая грудь, голубое брюшко), как то и ожидалось, кладки не различались ни по количеству, ни по размеру яиц. Однако обнаружено, что промежуточная морфа имеет более удлиненные яйца по сравнению с гетеро- и андроморфными самками.

© Виталий Сирота, Минский р-н
Замечено, что многочисленные тандемы, как правило, группами, занимают участки канав, покрытые Potamogeton natans. В 530 случаях (92,7% всех посадок) самка сразу же при приземлении изгибала брюшко и касалась листа яйцекладом. Контакт брюшка самки с растительным материалом длился от 2 сек до 12,47 мин., но не каждое посещение субстрата приводило к откладке яиц. Яйца откладывались почти исключительно в черешок и мясистое основание листа. При медленном движении самки и погружении брюшка в воду использовался черешок по всей его длине. Если тандему не мешали, яйцекладка продолжалась 3,72±2,47 мин и откладывалось 25,2±17,3 яиц. Яйца откладывались со средней скоростью 7 яиц/мин (4-13 яйца/мин). Скорость откладки яиц не коррелировала ни с температурой воды, ни с температурой воздуха. Кратковременные пребывания на листе без фактической откладки яиц имели среднюю продолжительность 16 сек (максимум 40 сек).

© Владимир Бондарь, Могилевский р-н
Во время посткопуляции тандемы проявляют агрегационное поведение. Группы развиваются, потому что тандемы предпочитают приземляться в местах, где уже присутствуют пары, откладывающие яйца. Особенно привлекательны тандемы с вертикально расположенным самцом. Можно считать, что самец-охранник обеспечивает фактический стимул.
Для откладывания яиц пара приземляется на конструкции, плавающие в воде или выступающие над поверхностью воды. На выбор места откладки яиц влияет наличие конспецификов и наличие определенных растительных структур. Вид предпочитает откладывать яйца во влажные или погруженные в воду части растений. Избегает восковой верхней поверхности листа и других несмачиваемых частей растения. В Potamogeton natans яйца откладываются почти исключительно в черешок. Тандемы преимущественно приземляются на основание черешка, даже когда он погружен в воду. Но это не всегда так. Часто тандемы ведут себя оппортунистически, приземляясь на пластину листа. Затем самка некоторое время ищет черешок брюшком. Если ей это не удается, после непродолжительного полета она ищет другую часть листа, пока в конце концов не находит черешок. Эта модель поведения, вероятно, особенно полезна при густой растительности и большом количестве черешков. Таким образом можно найти подходящие места для откладки яиц, которые невозможно обнаружить в полете.
Для откладывания яиц пара приземляется на конструкции, плавающие в воде или выступающие над поверхностью воды. На выбор места откладки яиц влияет наличие конспецификов и наличие определенных растительных структур. Вид предпочитает откладывать яйца во влажные или погруженные в воду части растений. Избегает восковой верхней поверхности листа и других несмачиваемых частей растения. В Potamogeton natans яйца откладываются почти исключительно в черешок. Тандемы преимущественно приземляются на основание черешка, даже когда он погружен в воду. Но это не всегда так. Часто тандемы ведут себя оппортунистически, приземляясь на пластину листа. Затем самка некоторое время ищет черешок брюшком. Если ей это не удается, после непродолжительного полета она ищет другую часть листа, пока в конце концов не находит черешок. Эта модель поведения, вероятно, особенно полезна при густой растительности и большом количестве черешков. Таким образом можно найти подходящие места для откладки яиц, которые невозможно обнаружить в полете.

♀ Фото © Владимир Бондарь, Могилевский р-н
Самка откладывает практически все выработанные ею созревшие яйца. Когда яйца были отложены накануне, среднее количество отложенных яиц на следующий день составило 191. Количество яиц увеличивается с увеличением количества дней, прошедших с момента последней откладки яиц. Было обнаружено, что максимальное количество яиц, откладываемых за один день, составляет около 400 с интервалом в 5 дней с момента предыдущей кладки. Среднее количество созревших яиц у вида не указывается, поскольку частота яйцекладки зависит от погоды и, таким образом, подвержена сильным колебаниям. Предполагая производство всего 191 яйца в день, принимая во внимание среднее количество яиц, отложенных при каждой кладке равное 25 яйцам в кладке, становится ясно, что пара должна откладывать яйца в несколько разных периодов. Средний период в тандеме составляет около 86 минут, хотя количество этого времени, затрачиваемого на фактическую откладку яиц, относительно невелико: для откладки 191 яйца со скоростью 7 яиц в минуту тандему потребуется всего 27 мин. Значительная часть оставшегося времени уходит на поиск подходящих мест для кладки.

Фото © Владимир Бондарь, Могилевский р-н
Вертикальная защитная позиция самцов известна давно, но значение этой позы все еще плохо изучено. Предлагаются различные объяснения. Во-первых, это снижает риск нападения хищников. При нападениях зеленых лягушек тандему легче избежать их, когда самец находится в вертикальном положении. Это позволяет им дольше оставаться в пределах досягаемости приближающегося хищника. Во-вторых, поза стимулирует формирование скоплений яйцекладущих тандемов. Вертикальное расположение тандемного самца оказывает гораздо более сильное притягательное воздействие на другие тандемы, чем горизонтальное, а яйцекладка в группах имеет много положительных эффектов, один из которых - тревога, поднимаемая всей группой при нападении хищника на один из тандемов, которая позволяет скрыться остальным парам.
Кроме того, поза самца позволяет тандему приземлиться на более мелкие кусочки субстрата, поскольку не требуется места для самца. Можно приземляться на верхушки растений, которые в противном случае были бы погружены в воду. Всё это значительно увеличивает выбор потенциальных мест для откладки яиц.
Гипотеза, что поза предотвращает перегрев тела самца выглядит сомнительно, по крайней мере, для этого вида. Также как и вариант того, что вертикальное положение может обеспечить лучшую защиту самки от соперничающих самцов, пытающихся разделить тандем. У C. puella, хотя и наблюдались агрессивные нападения на тандемы, это никогда не приводило к их разъединению во время откладки яиц.
Кроме того, поза самца позволяет тандему приземлиться на более мелкие кусочки субстрата, поскольку не требуется места для самца. Можно приземляться на верхушки растений, которые в противном случае были бы погружены в воду. Всё это значительно увеличивает выбор потенциальных мест для откладки яиц.
Гипотеза, что поза предотвращает перегрев тела самца выглядит сомнительно, по крайней мере, для этого вида. Также как и вариант того, что вертикальное положение может обеспечить лучшую защиту самки от соперничающих самцов, пытающихся разделить тандем. У C. puella, хотя и наблюдались агрессивные нападения на тандемы, это никогда не приводило к их разъединению во время откладки яиц.

♂ Фото © Владимир Бондарь, Могилевский р-н
Нимфа. Окраска тела варьирует: желтоватая, желтовато-зеленая, зеленоватая, коричневатая с красновато-коричневыми отметинами. Тело длинное, стройное и гладкое. Длина 21-22 мм, ширина 2,5-3 мм. Голова широкая, около 4 мм в поперечнике, от лба до заднего края затылка шириной около 2 мм, с более светлыми пятнами на передней части; глаза большие, почти эллипсоидной формы, темно оливково-коричневые, почти не заходящие или очень мало заходящие за внешние края затылка; затылок относительно широкий, впереди округлый, внешние края скошены, задние углы округлые, не выдающиеся, обрамлены шипиками, задний край глубоко вогнутый; вся поверхность затылка покрыта красновато-коричневыми точками. Маска в состоянии покоя достигает основания передней пары ног; подбородок узкий у основания, довольно широкий у вершины. Переднегрудь большая, почти пятиугольная, с темными краями: дыхальца среднегруди хорошо видимые, темные; грудь покрыта красновато-коричневыми точками. Крыловые чехлы короткие, приблизительно 4 мм, доходят до середины IV сегмента брюшка или немного дальше. Ноги светлые, с дистальным темным кольцом на бедрах и иногда на голенях, где оно большею частью едва заметно; иногда на бедрах имеется еще одно, несколько более бледное базальное кольцо; дистальные концы лапок и места сочленений между члениками ног также темные; ноги вооружены рядами красновато-темных шипов. Брюшко цилиндрическое, постепенно суживающееся к концу; на всех сегментах до VIII включительно проходят дорсально 3 светлые продольные полосы: 2 по бокам, а 1 посредине, с каждой стороны последней темная красно-коричневая полоска, более широкая у переднего края сегмента и постепенно суживающаяся к заднему краю; все брюшко, помимо указанных полос, покрыто красновато-коричневыми точками; межсегментальные пространства, за исключением Х сегмента, с кольцом из темных точек; задний край Х сегмента усеян жесткими волосками; боковые края сегментов, вооружены короткими шипиками. Хвостовые жаберные пластинкя довольно длинные, 5-6,5 мм, шириной 1,5 мм, ланцетовидные, тупо заостренные на вершине, охристо-коричневые, главный средний трахеальный ствол и разветвления трахей коричневые; поперечный узелковый шов косой, узелки расположены также наискось по отношению друг к другу; оба края дистальной части обрамлены волосками; разветвление трахей густое и четкое; в области поперечного узелкового шва проходит темная поперечная полоса. Церкоиды небольшие, малозаметные; створки яйцеклада доходят до середины Х тергита.

© Екатерина Морозова (Букас), Ивацевичский р-н, Брестская обл.
Экология вида во многом тождественна с экологией С. pulcliellum, так как оба вида почти всегда встречаются одновременно. Личинки заселяют самые разнообразные водоемы: речные заводи, старицы, озера, пруды, осоковые и торфяные болота, лужи, но предпочитают местообитания с богатой водной растительностью. В отличие от C. pulchellum, в среднем достигает более высокой численности в свободных от рыбы водоемах, но при оптимально разнообразной структурой растительности, могут заселяться и водоемы с хорошо развитой ихтиофауной.
Личинки вылупляются перед зимой и зимуют в основном на стадии 4-го возраста. Развитие личинок завершается за 4 недели. В то же время сроки развития различаются географически. Например, для Британии и Ирландии указывается, что полное развитие от яйца до имаго, как и во всей Европе, обычно занимает 1 год, но может занять и 2 года.
Появление крылатых особей происходит утром на надводной растительности, включая соцветия рдеста в центре пруда, при этом самки появляются на день или два раньше, чем самцы, и с меньшей вероятностью, чем самцы, вернутся в места своего отрождения.
Отрождения имаго начинаются в мае. Если погода солнечная, строго синхронизированная основная часть отрождений происходит через 2-3 недели после появления первых имаго. Затем скопления только что вылупившихся и достигающих зрелости имаго обнаруживаются на защищенных от ветра открытых пространствах вблизи воды. Отрождения значительно замедляется в начале июня и часто заканчиваются в его середине, но в отдельные годы отдельные только что отродившиеся особи регистрировались в конце июля или начале августа.
Личинки вылупляются перед зимой и зимуют в основном на стадии 4-го возраста. Развитие личинок завершается за 4 недели. В то же время сроки развития различаются географически. Например, для Британии и Ирландии указывается, что полное развитие от яйца до имаго, как и во всей Европе, обычно занимает 1 год, но может занять и 2 года.
Появление крылатых особей происходит утром на надводной растительности, включая соцветия рдеста в центре пруда, при этом самки появляются на день или два раньше, чем самцы, и с меньшей вероятностью, чем самцы, вернутся в места своего отрождения.
Отрождения имаго начинаются в мае. Если погода солнечная, строго синхронизированная основная часть отрождений происходит через 2-3 недели после появления первых имаго. Затем скопления только что вылупившихся и достигающих зрелости имаго обнаруживаются на защищенных от ветра открытых пространствах вблизи воды. Отрождения значительно замедляется в начале июня и часто заканчиваются в его середине, но в отдельные годы отдельные только что отродившиеся особи регистрировались в конце июля или начале августа.

Фото © Владимир Бондарь, Могилевский р-н

♂ Фото © Семён Левый, Шарковщинский р-н, Витебская обл.

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
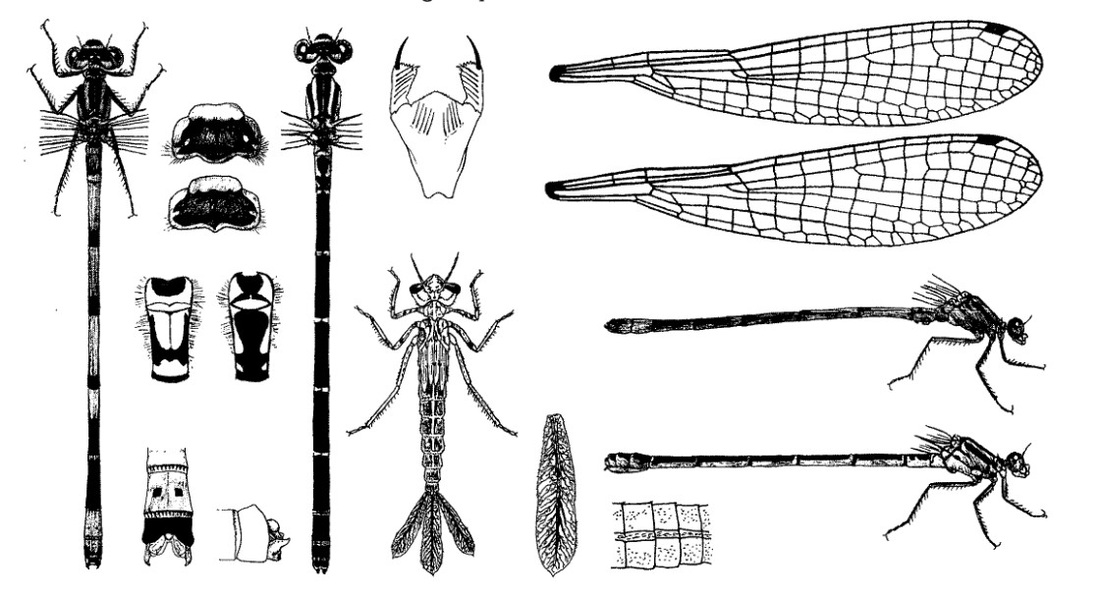
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Azure Bluet. Coenagrion puella" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 70-71.
9. Smallshire D., Swash A. "Azure Damselfly. Coenagrion puella" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 86-87.
10. Martens A. "Field experiments on aggregation behaviour and oviposition in Coenagrion puella (L.) (Zygoptera: Coenagrionidae)" / Advances in Odonatololgy, 6 : 49-58. 1994.
11. Joop G., Gillen A., Mikolajewski D. J. "Colour polymorphism in female Coenagrion puella: differences in egg shape (Odonata: Coenagrionidae)" / International Journal of Odonatology 10 (1) 2007: 71-80.
12. Martín R. "Coenagrion pulchellum (Van der Linden, 1825) (Insecta, Odonata, Coenagrionidae) en la Península Ibérica" / Boletín de la Sociedad Entomológica Aragonesa (S.E.A.), nº 48 (30/06/2011): 493-495.
13. Lehmann A., Lehmann G., Petzold F., Mauersberger R. "Hufeisen-Azurjungfer – Coenagrion puella (Linnaeus 1758)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 44-45.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
6. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Azure Bluet. Coenagrion puella" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 70-71.
9. Smallshire D., Swash A. "Azure Damselfly. Coenagrion puella" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 86-87.
10. Martens A. "Field experiments on aggregation behaviour and oviposition in Coenagrion puella (L.) (Zygoptera: Coenagrionidae)" / Advances in Odonatololgy, 6 : 49-58. 1994.
11. Joop G., Gillen A., Mikolajewski D. J. "Colour polymorphism in female Coenagrion puella: differences in egg shape (Odonata: Coenagrionidae)" / International Journal of Odonatology 10 (1) 2007: 71-80.
12. Martín R. "Coenagrion pulchellum (Van der Linden, 1825) (Insecta, Odonata, Coenagrionidae) en la Península Ibérica" / Boletín de la Sociedad Entomológica Aragonesa (S.E.A.), nº 48 (30/06/2011): 493-495.
13. Lehmann A., Lehmann G., Petzold F., Mauersberger R. "Hufeisen-Azurjungfer – Coenagrion puella (Linnaeus 1758)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 44-45.
