СТРЕКОЗА ДЛИННОХВОСТАЯ
Leucorrhinia caudalis Charpentier, 1840

♂ Фото © Семён Левый, Смолевичский р-н, Минская обл.
Leucorrhinia caudalis Charpentier, 1840 (стрекоза длиннохвостая)
Нижняя губа вся черная. Лоб кремово-белый, глаза темные. Тело в основном черное. Брюшко короткое и широкое. VI-VIII сегменты брюшка сильно веретенообразно расширены, особенно у самца. Анальные придатки, хотя бы частично белые. Основание задних крыльев с темным непрозрачным пятном. Передние крылья без темного базального пятна, задние - с пятном. Жилкование крыльев темное. Ноги темные. Придатки белые.
♂: Грудь черная с неясными или практически незаметными предплечевыми полосами и желтоватыми пятнышками на боках. Брюшко черное, II-III сегменты по бокам с желтым пятном; III-VI(V) сегменты с голубым налетом. У взрослых птеростигма белая, окруженная толстыми черными жилками, от птеростигмы к кончику крыла расходятся белые жилки (не всегда заметные).
♀: Предплечевые полосы более ясные, оранжево-желтые, боковые пятна неправильной формы на груди и желтые отметки между основаниями крыльев ярче и больше. Брюшко с желтоватыми пятнами по середине II-VI сегментов; сегменты II-IV с боковыми желтыми пятнышками. На основаниях крыльев желтые пятна, у некоторых самок за каждым пятном на крыле есть нечеткое темное пятно. Птеростигма буровато-черная, от нее к кончику крыла расходятся белые жилки. Генитальная пластинка самки большая, до половины IX сегмента, глубоко расщепленная.
Неполовозрелые особи черно-желтые, как самки.
Этот вид и L. albifrons представляют собой два европейских вида, у которых самцы темные, с голубым налетом на его основании и темным в остальном брюшком, на котором отсутствуют красные отметки (от последнего отличается более контрастно выраженным голубым налетом). От L. albifrons отличается явно вздутыми сегментами VII-VIII, у самцов в основании брюшка. У самок желтые пятна по бокам груди и желтые отметки между основаниями крыльев больше, чем у L. albifrons.
Лет: май - июль (главным образом май и июнь).
Длина 33-37 мм, брюшко 23-24 мм, заднее крыло 29-32 мм.
♂: Грудь черная с неясными или практически незаметными предплечевыми полосами и желтоватыми пятнышками на боках. Брюшко черное, II-III сегменты по бокам с желтым пятном; III-VI(V) сегменты с голубым налетом. У взрослых птеростигма белая, окруженная толстыми черными жилками, от птеростигмы к кончику крыла расходятся белые жилки (не всегда заметные).
♀: Предплечевые полосы более ясные, оранжево-желтые, боковые пятна неправильной формы на груди и желтые отметки между основаниями крыльев ярче и больше. Брюшко с желтоватыми пятнами по середине II-VI сегментов; сегменты II-IV с боковыми желтыми пятнышками. На основаниях крыльев желтые пятна, у некоторых самок за каждым пятном на крыле есть нечеткое темное пятно. Птеростигма буровато-черная, от нее к кончику крыла расходятся белые жилки. Генитальная пластинка самки большая, до половины IX сегмента, глубоко расщепленная.
Неполовозрелые особи черно-желтые, как самки.
Этот вид и L. albifrons представляют собой два европейских вида, у которых самцы темные, с голубым налетом на его основании и темным в остальном брюшком, на котором отсутствуют красные отметки (от последнего отличается более контрастно выраженным голубым налетом). От L. albifrons отличается явно вздутыми сегментами VII-VIII, у самцов в основании брюшка. У самок желтые пятна по бокам груди и желтые отметки между основаниями крыльев больше, чем у L. albifrons.
Лет: май - июль (главным образом май и июнь).
Длина 33-37 мм, брюшко 23-24 мм, заднее крыло 29-32 мм.

♂ Фото © Виктор Рындевич, Смолевичский р-н, Минская обл.
Населяет теплые, хорошо освещаемые солнцем, густо заросшие озера, пруды и старицы, обычно в лесных ландшафтах. Встречается и на участках с прозрачными и хорошо насыщенными кислородом водами крупных рек, а также на старых карьерах мезотрофных торфяников. Указываются в том числе и искусственные водоемы, с глубокой чистой водой, обильной подводной и часто плавающей растительностью. Лучше других видов рода переносит присутствие рыб.
Благоприятные водоемы встречаются в различных типах более или менее лесистой среды - важно наличие поблизости деревьев или кустарников. В лесу участок должен оставаться достаточно солнечным. Однако в некоторых швейцарских стациях, видимо, наличие древесных растений не является строгим требованием, потому что большие популяции присутствуют на участках, где их нет.
Подавляющее большинство стаций относятся к водоемам антропогенного происхождения. Часто это бывшие участки добычи торфа или рыбные пруды. Вид также колонизирует заводи и старые карьеры с хорошей растительностью. Наиболее благоприятные водоемы достаточно большие (не менее 0,5 га), питаются подземными водами и не имеют притоков. Однако это нельзя обобщать. Пруды площадью 30 м² также могут быть благоприятными.
Благоприятные водоемы встречаются в различных типах более или менее лесистой среды - важно наличие поблизости деревьев или кустарников. В лесу участок должен оставаться достаточно солнечным. Однако в некоторых швейцарских стациях, видимо, наличие древесных растений не является строгим требованием, потому что большие популяции присутствуют на участках, где их нет.
Подавляющее большинство стаций относятся к водоемам антропогенного происхождения. Часто это бывшие участки добычи торфа или рыбные пруды. Вид также колонизирует заводи и старые карьеры с хорошей растительностью. Наиболее благоприятные водоемы достаточно большие (не менее 0,5 га), питаются подземными водами и не имеют притоков. Однако это нельзя обобщать. Пруды площадью 30 м² также могут быть благоприятными.

♀ Фото Владимир Бондарь, Быховский р-н, Могилевская обл.
Водная растительность является ключевым элементом. Наличие кувшинок или водорослей - очень благоприятный фактор. Оптимальным является покрытие плавающими растениями от 30 до 70%. Однако чрезмерное покрытие поверхности водоема также неблагоприятно. Очень важно для развития личинок наличие подводной растительности (в частности, харовых водорослей, перистолистника, пузырчатки или водного лютика). Покрытие от 10 до 50% в зависимости от типа растительности кажется оптимальным. Пруды без растительности или слишком заросшие - неблагоприятны.
Глубина занимаемых водоемов непостоянна. В районе исследования в самых глубоких водоемах (от 1,5 до 2 м по всей их поверхности) вид отсутствовал. Авторы исследования связывают это с тем, что растительность не может развиваться на слишком большой глубине. С другой стороны, в Швейцарии идеальные водоемы имеют глубину от 1,5 до 3 м (максимум 5 м).
Пологие берега обычно считаются более благоприятными, но вид встречается на прудах с крутыми берегами.
Глубина занимаемых водоемов непостоянна. В районе исследования в самых глубоких водоемах (от 1,5 до 2 м по всей их поверхности) вид отсутствовал. Авторы исследования связывают это с тем, что растительность не может развиваться на слишком большой глубине. С другой стороны, в Швейцарии идеальные водоемы имеют глубину от 1,5 до 3 м (максимум 5 м).
Пологие берега обычно считаются более благоприятными, но вид встречается на прудах с крутыми берегами.

♂ Фото © Виктор Рындевич, Смолевичский р-н, Минская обл.
Во время фазы созревания молодые особи могут удаляться от мест размножения, и их можно наблюдать в различных средах, таких как болота или пустыри, а также на влажных лугах у водоема. Затем зрелые взрослые встречаются в основном над водоемом, иногда на значительной высоте, а также среди ветвей близлежащих деревьев и кустарников, особенно те самцы, которые ожидают появление свободных территорий на водоеме или в пасмурную погоду.
Оба пола часто приземляются на плавающую растительность, особенно на листья кувшинок, где самцы устанавливают свою территорию и высматривают возможную добычу. Однако если самцы обычно остаются в непосредственной близости от мест размножения, а самки прилетают туда только для спаривания и откладывания яиц. Передвижения по участку в основном связаны с защитой территории у самцов и поиском места для кладки у самок.
В пределах участка расстояние перемещения, вероятно, зависит от размера водоема и окружающей среды.
Активность меняется в течение дня: она максимальна с конца утра до начала дня. Кроме того, в масштабе участка на передвижения влияет ветер, когда защищенные участки посещаются гораздо чаще. При этом наличие кустарников или крупных гелофитов на берегах является важным параметром приживаемости вида. Вероятно, вид не особенно склонен к экспансии. Однако он вполне способен быстро колонизировать участки на расстоянии от 5 до 10 км, отмечены расселения и до 50 км.
Оба пола часто приземляются на плавающую растительность, особенно на листья кувшинок, где самцы устанавливают свою территорию и высматривают возможную добычу. Однако если самцы обычно остаются в непосредственной близости от мест размножения, а самки прилетают туда только для спаривания и откладывания яиц. Передвижения по участку в основном связаны с защитой территории у самцов и поиском места для кладки у самок.
В пределах участка расстояние перемещения, вероятно, зависит от размера водоема и окружающей среды.
Активность меняется в течение дня: она максимальна с конца утра до начала дня. Кроме того, в масштабе участка на передвижения влияет ветер, когда защищенные участки посещаются гораздо чаще. При этом наличие кустарников или крупных гелофитов на берегах является важным параметром приживаемости вида. Вероятно, вид не особенно склонен к экспансии. Однако он вполне способен быстро колонизировать участки на расстоянии от 5 до 10 км, отмечены расселения и до 50 км.

♀ Фото Владимир Бондарь, Быховский р-н, Могилевская обл.
На границах ареала более или менее изолированные популяции очень небольшие. Однако наблюдаются и более крупные. Так, в Роне-Альпах наибольшее количество особей, наблюдаемых одновременно на водоеме, составляет более 50 самцов, но следует учитывать территориальность вида: многие самцы остаются вдали от воды. Популяции Лотарингии часто составляют от 250 до 500 наблюдаемых особей, а плотность экзувий составляет 120 на 50 м береговой линии.
Исследования в Швейцарии показало большую изменчивость численности популяции: от менее 20 до 9000 особей. Отмечается, что участки с наименьшей популяцией не заселяются постоянно. Кроме того, у вида часто наблюдаются сильные колебания численности, что может быть связано с погодными условиями во время окрыления имаго.
Отсутствует информация о минимальной площади, при которой популяция может быть жизнеспособной. Однако вид может развиваться в небольших прудах: размер водоема колеблется от 30 м² до 220 га, чаще всего от 0,4 до 2 га.
Считается, что популяция находится в отличном состоянии, если количество взрослых самцов, наблюдаемых в водоеме, превышает 30 особей. Состояние считается плохим при количестве ниже 5 самцов. Предложена также оценка по экзувиям: сумма двух посещений в период отрождения должна позволять найти не менее 1 экзувия на 1 м берега для популяции в «отличном» состоянии, и в «плохом», если на 1 м приходится менее 0,1 экзувия.
Исследования в Швейцарии показало большую изменчивость численности популяции: от менее 20 до 9000 особей. Отмечается, что участки с наименьшей популяцией не заселяются постоянно. Кроме того, у вида часто наблюдаются сильные колебания численности, что может быть связано с погодными условиями во время окрыления имаго.
Отсутствует информация о минимальной площади, при которой популяция может быть жизнеспособной. Однако вид может развиваться в небольших прудах: размер водоема колеблется от 30 м² до 220 га, чаще всего от 0,4 до 2 га.
Считается, что популяция находится в отличном состоянии, если количество взрослых самцов, наблюдаемых в водоеме, превышает 30 особей. Состояние считается плохим при количестве ниже 5 самцов. Предложена также оценка по экзувиям: сумма двух посещений в период отрождения должна позволять найти не менее 1 экзувия на 1 м берега для популяции в «отличном» состоянии, и в «плохом», если на 1 м приходится менее 0,1 экзувия.

♀ Фото Владимир Бондарь, Быховский р-н, Могилевская обл.
Самцы территориальны и проявляют явную агрессивность по отношению к своим сородичам или другим стрекозам. Они защищают территорию от 10 до 20 м² со своего наблюдательного пункта (особенно с листьев кувшинок), часто сидя с приподнятым брюшком. В отличие от других видов рода, их территориальность сохраняется, а агрессивность по отношению к соперникам возрастает с увеличением плотности популяции. Спаривание, которое длится от 5 до 30 минут, происходит на территории самца.
Самка начинает откладывать яйца на территории самца, затем выходит за ее пределы. Самку может охранять самец, который затем улетает к ближайшей фиксированной точке своей территории или откладка яиц может совершаться в одиночку. Яйца откладываются в полете, в плавающую или подводную растительность. Откладка наблюдалась на скопления перистолистника и рдеста, а также роголистника. При кладке яиц самка летает большими кругами над поверхностью воды и время от времени опускает кончик брюшка в воду, выбрасывая яйца; часто она так сильно ударяет кончиком брюшка о поверхность воды, что вверх летят брызги воды.
Самка начинает откладывать яйца на территории самца, затем выходит за ее пределы. Самку может охранять самец, который затем улетает к ближайшей фиксированной точке своей территории или откладка яиц может совершаться в одиночку. Яйца откладываются в полете, в плавающую или подводную растительность. Откладка наблюдалась на скопления перистолистника и рдеста, а также роголистника. При кладке яиц самка летает большими кругами над поверхностью воды и время от времени опускает кончик брюшка в воду, выбрасывая яйца; часто она так сильно ударяет кончиком брюшка о поверхность воды, что вверх летят брызги воды.

Фото © Владимир Бондарь, Быховский р-н, Могилевская обл.
Нимфа с широким коротким пестро разрисованным телом. Голова маленькая, широкая, короткая, длина почти в 2 раза короче ширины; темя приподнятое; затылок очень хорошо развитый, посредине укороченный, с округлыми задними углами и вогнутым задним краем. На задних крыловых чехлах у основания находятся светлые ячейки, окруженные темными гранями. Ноги длинные, бедра с 2 темными кольцами и несколькими темными точками. Брюшко овальной формы, короткое, широкое, постепенно расширяющееся до VI сегмента включительно и затем постепенно суживающееся к IX сегменту; с дорсальной стороны с темными пятнами, расположенными в 3 ряда, из которых средние пятна самые темные и самые резкие, по бокам ряд светлых пятен; с вентральной стороны 2 ряда темных пятен, соединяющихся на последних сегментах в поперечные полосы; Х сегмент маленький, узкий. Дорсальный придаток анальной пирамиды короче приблизительно на 1/6 церков, сильно расширенный у основания, с острой тонкой вершиной; церки по длине приблизительно равны IX и Х сегментам, взятым вместе, с острой вершиной; церкоиды в 2 раза короче церков, заостренные на вершине.

♀ Фото Владимир Бондарь, Быховский р-н, Могилевская обл.
Личинки обитают в водоемах с большой и открытой водной поверхностью, у берегов, густо заросших водными растениями: Nymphaea candida, Nuphar luteum, Scirpus lacustris, Polygonum amphibium и др. К таким водоемам можно отнести довольно глубокие с сравнительно чистой водой пруды и подобные им водоемы (заливы озер, заводи и бухты рек). Личинки имеют хорошо развитые боковые и спинные шипы на различных сегментах брюшка, что в 4 раза снижает риск хищничества от нападения рыб сзади. Таким образом, они могут противостоять присутствию некоторых видов рыб, тем более что прячутся среди подводных зарослей водорослей, которые служат им убежищем. Мало того, сосуществование личинок с кормовой цепью, контролируемой рыбой, показало оптимальное заселение водоемов с преобладанием плотвы и окуня, часто в сочетании с многочисленными другими видами рыб. Причина заключается в том, что личинки довольно хорошо защищены от хищничества рыб, но чувствительны к беспозвоночным хищникам, таким как, например, личинки стрекоз или жуки-плавунцы. Однако присутствие белого амура может нанести ущерб водной растительности и, следовательно, угрожать популяции личинок.
Передвижение личинок осуществляется по субстрату на дне водоема. Молодые личинки укрываются в подводной растительности в поверхностной части водоема. Подрастая, они могут вернуться глубже и прятаться в донных отложениях.
Фаза личинки обычно длится 2 года, если вода теплая, возможно завершение за 1 год.
Отрождения имаго почти во все годы начинаются во второй декаде мая. Отрождение происходит днем, преимущественно утром, на растительности на берегах у кромки воды или на мелководье, на высоте 5-30 см над ее поверхностью, реже до 60 см. Субстратами для отрождения являются растения с узким стеблем, особенно осока (Carex spp.). Обычно начинается с середины мая (в иные годы раньше или позже). Отрождение происходит в течение 4 недель, а созревание молодых особей длится около 10 дней.
Передвижение личинок осуществляется по субстрату на дне водоема. Молодые личинки укрываются в подводной растительности в поверхностной части водоема. Подрастая, они могут вернуться глубже и прятаться в донных отложениях.
Фаза личинки обычно длится 2 года, если вода теплая, возможно завершение за 1 год.
Отрождения имаго почти во все годы начинаются во второй декаде мая. Отрождение происходит днем, преимущественно утром, на растительности на берегах у кромки воды или на мелководье, на высоте 5-30 см над ее поверхностью, реже до 60 см. Субстратами для отрождения являются растения с узким стеблем, особенно осока (Carex spp.). Обычно начинается с середины мая (в иные годы раньше или позже). Отрождение происходит в течение 4 недель, а созревание молодых особей длится около 10 дней.

♀ Фото Владимир Бондарь, Быховский р-н, Могилевская обл.

♂ Фото © Владимир Бондарь, Могилевский р-н

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0

♀ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
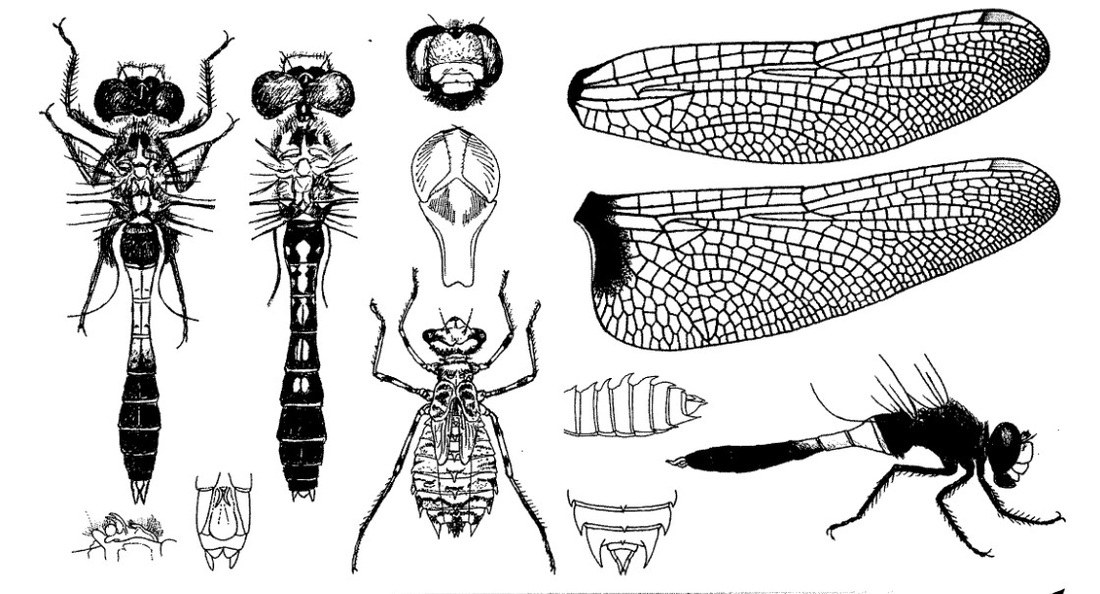
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Smallshire D., Swash A. "Lilypad Whiteface. Leucorrhinia caudalis" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 292-293.
8. Merlet F., Houard X. "Synthèse bibliographique sur les traits de vie de la Leucorrhine à large queue (Leucorrhinia caudalis (Charpentier, 1840)) relatifs à ses déplacements et à ses besoins de continuités écologiques". Office pour les insectes et leur environnement & Service du patrimoine naturel du Muséum national d’Histoire naturelle. Paris, 2012. 9 pages.
9. Vonwil G. "Fiches de protection espèces – Libellules – Leucorrhinia caudalis". Groupe de travail pour la conservation des Libellules de Suisse, CSCF info fauna, Neuchâtel et Office fédéral de l’environnement, Berne, 2013. 5 p.
10. Jacquot P. "Les libellules menacées en Franche-Comté. Leucorrhine à large queue Leucorrhinia caudalis (Charpentier, 1840). Union européenne, Direction Régionale de l’Environnement, de l’Aménagement et du Logement de Franche-Comté, Conseil régional de Franche-Comté, 2012. - 2 p.
11. Mauersberger R. "Zierliche Moosjungfer – Leucorrhinia caudalis (Charpentier 1840)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 112-113.
12. Бондарь В. А. "Leucorrhinia caudalis на мелководных водоемах, оставшихся после добычи торфа между деревнями Годылево и Перекладовичи Быховского р-на Могилевской обл." Личное сообщение с фотоматериалом от 26.05.2023.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1953. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Smallshire D., Swash A. "Lilypad Whiteface. Leucorrhinia caudalis" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 292-293.
8. Merlet F., Houard X. "Synthèse bibliographique sur les traits de vie de la Leucorrhine à large queue (Leucorrhinia caudalis (Charpentier, 1840)) relatifs à ses déplacements et à ses besoins de continuités écologiques". Office pour les insectes et leur environnement & Service du patrimoine naturel du Muséum national d’Histoire naturelle. Paris, 2012. 9 pages.
9. Vonwil G. "Fiches de protection espèces – Libellules – Leucorrhinia caudalis". Groupe de travail pour la conservation des Libellules de Suisse, CSCF info fauna, Neuchâtel et Office fédéral de l’environnement, Berne, 2013. 5 p.
10. Jacquot P. "Les libellules menacées en Franche-Comté. Leucorrhine à large queue Leucorrhinia caudalis (Charpentier, 1840). Union européenne, Direction Régionale de l’Environnement, de l’Aménagement et du Logement de Franche-Comté, Conseil régional de Franche-Comté, 2012. - 2 p.
11. Mauersberger R. "Zierliche Moosjungfer – Leucorrhinia caudalis (Charpentier 1840)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 112-113.
12. Бондарь В. А. "Leucorrhinia caudalis на мелководных водоемах, оставшихся после добычи торфа между деревнями Годылево и Перекладовичи Быховского р-на Могилевской обл." Личное сообщение с фотоматериалом от 26.05.2023.
