ОГНЕТЕЛКА НИМФАЛЬНАЯ
Pyrrhosoma nymphula Sulzer, 1776

♂ Фото © Семён Левый, г. Могилев
Pyrrhosoma nymphula Sulzer, 1776 (огнетелка нимфальная или краснотелка-нимфа)
Голова широкая, сверху вдвое шире своей длины. Затылок без светлых пятен. Глаза красно-коричневые, пересекаемые двумя темными горизонтальными полосами, у самок более тусклые. Задний край переднеспинки равномерно закруглен. Основной тон окраски красный. Верх груди и ее бока в передней части бронзово-черные, металлически-блестящие, под метаплевральным швом желтая цвет с черной точечной маркировкой. Светлоокрашенная предплечевая полоса полная, широкая, пересекающая плечевой шов. Ниже ее по груди проходит широкая черная полоса. Сложенные крылья заходят за VII сегмент брюшка. Крылья прозрачные. Птеростигма узкая (равна 1 ячейке), темная. Ноги черные.
♂: Предплечевые полосы широкие красные. Брюшко темно-красное с бронзово-черными дорсальными полосами на сегментах VII-IX, которые могут быть плохо заметны при ярком солнечном свете. На сочленениях сегментов узкие черные кольца. Если смотреть сбоку, нижние придатки незначительно короче верхних, с «крючком», выступающим вверх у кончика, примерно на две трети длины верхних.
♀: Задний край переднеспинки с двумя небольшими складками, не приподнятыми при виде сбоку. Предплечевые полосы широкие желтые, в зависимости от формы разной интенсивности (от ярких до бледных, почти белых). Имеет три цветовые формы, каждая из которых имеет узкие желтые кольца на переднем конце II-VII сегментов брюшка. Форма typica (intermedia), похожа на самца, но имеет ярко-желтые предплечевые полосы и более обширный черный цвет на всех сегментах брюшка, черная полоса на VI сегменте брюшка проходит по всей длине сегмента, а на заднем конце первых пяти сегментов брюшка имеются черные полосы. У темной формы melanotum бледно-желтые предплечевые полосы (как и у неполовозрелых особей обоих полов) и очень ограниченные участки красного и желтого цвета на большей части черного брюшка; также глаза более тускло-красные. У некоторых особей этой формы красный цвет на брюшке сменяется желтым, хотя такие особи, возможно, и не достигли своей окончательной окраски. У красной формы fulvipes черный цвет на брюшке занимает промежуточное положение между typica и самцом, похожа по окраске на самца, но также имеет черные полосы на заднем конце V и VI сегментов брюшка, а черный цвет на VII и VIII сегментах более обширен, чем у самца. У всех форм имеются узкие желтые полосы между сегментами, которые становятся красными к кончику.
У неполовозрелых особей бледно-желтые плечевые полосы, глаза тусклые.
Длина 33-36 мм, брюшко 25-27 мм, заднее крыло 21-23 мм.
Лет: май - конец августа.
♂: Предплечевые полосы широкие красные. Брюшко темно-красное с бронзово-черными дорсальными полосами на сегментах VII-IX, которые могут быть плохо заметны при ярком солнечном свете. На сочленениях сегментов узкие черные кольца. Если смотреть сбоку, нижние придатки незначительно короче верхних, с «крючком», выступающим вверх у кончика, примерно на две трети длины верхних.
♀: Задний край переднеспинки с двумя небольшими складками, не приподнятыми при виде сбоку. Предплечевые полосы широкие желтые, в зависимости от формы разной интенсивности (от ярких до бледных, почти белых). Имеет три цветовые формы, каждая из которых имеет узкие желтые кольца на переднем конце II-VII сегментов брюшка. Форма typica (intermedia), похожа на самца, но имеет ярко-желтые предплечевые полосы и более обширный черный цвет на всех сегментах брюшка, черная полоса на VI сегменте брюшка проходит по всей длине сегмента, а на заднем конце первых пяти сегментов брюшка имеются черные полосы. У темной формы melanotum бледно-желтые предплечевые полосы (как и у неполовозрелых особей обоих полов) и очень ограниченные участки красного и желтого цвета на большей части черного брюшка; также глаза более тускло-красные. У некоторых особей этой формы красный цвет на брюшке сменяется желтым, хотя такие особи, возможно, и не достигли своей окончательной окраски. У красной формы fulvipes черный цвет на брюшке занимает промежуточное положение между typica и самцом, похожа по окраске на самца, но также имеет черные полосы на заднем конце V и VI сегментов брюшка, а черный цвет на VII и VIII сегментах более обширен, чем у самца. У всех форм имеются узкие желтые полосы между сегментами, которые становятся красными к кончику.
У неполовозрелых особей бледно-желтые плечевые полосы, глаза тусклые.
Длина 33-36 мм, брюшко 25-27 мм, заднее крыло 21-23 мм.
Лет: май - конец августа.

♀ Фото © Виктор Рындевич, Минский р-н
Встречается в очень широком диапазоне мест обитания, и в стоячих, и в проточных водоемах, однако везде предпочитает хорошо развитую водную и прибрежную растительность. Однако локально места обитания несколько разнятся. Для Восточной Европы, указывается, что вид заселяет преимущественно медленно текущие речки и ручьи, берега которых густо поросли кустарниками. Для Западной Европы указывается большое разнообразие водно-болотных угодий, от кислых болотных луж до солоноватых канав, особенно защищенные водоемы с обильными водными растениями. Вместе с тем, для Испании указываются среднее и нижнее течение рек и ручьев, искусственные пруды и резервуары для воды; в основном встречаются в слабо проточных или стоячих водоемах, богатых прибрежной растительностью, такой как тростник и камыш.
В Бранденбурге (Германия) только 2% регистраций получены из небольших водоемов, а около 40% составляют проточные воды, особенно медленно текущие ручьи, канавы и каналы. Хотя вид широко распространен на озерах, его численность там обычно ниже средней. С другой стороны, и пруды, и кислые болотные озера заселяются с высокой степенью непрерывности. Также поразительно значительное содержание гумуса во многих местообитаниях; оптимальными кажутся воды не слишком мутные и быстро прогревающиеся из-за темного цвета. Водоемы очень часто окружены деревьями, которые обеспечивают защиту от ветра.
В Бранденбурге (Германия) только 2% регистраций получены из небольших водоемов, а около 40% составляют проточные воды, особенно медленно текущие ручьи, канавы и каналы. Хотя вид широко распространен на озерах, его численность там обычно ниже средней. С другой стороны, и пруды, и кислые болотные озера заселяются с высокой степенью непрерывности. Также поразительно значительное содержание гумуса во многих местообитаниях; оптимальными кажутся воды не слишком мутные и быстро прогревающиеся из-за темного цвета. Водоемы очень часто окружены деревьями, которые обеспечивают защиту от ветра.

♀ Фото © Виктор Рындевич, Минский р-н
Самцы появляются немного раньше и в большем количестве, чем самки, созревают примерно за 12 дней (6-15 дней), что на 4 дня меньше, чем у самок. Впрочем, для второй, несинхронной летней волны отрождений указывается, что созревание самцов занимает около 6 дней. Репродуктивно зрелые взрослые особи могут жить в среднем около недели, но некоторые доживают до 35-45 дней.
Согласно другим исследованиям, средняя продолжительность жизни половозрелых особей, т. е. тех, кто пережил период созревания, составляет 6,8 дня для самцов и 6,6 дня для самок. Эти цифры дают общую продолжительность имаго 19,4 дня (12,6 предрепродуктивных + 6,8 репродуктивных) для самцов и 21,6 дней (15 предрепродуктивных + 6,6 репродуктивных) для самок. Вместе с тем максимальная зарегистрированная продолжительность жизни имаго - 46 дней.
Летают между растениями, растущими близко к берегам рек, между камышами и тростниками. В жаркую погоду летают быстро. В холодную погоду могут греться на светлых поверхностях, включая одежду. Отдыхают горизонтально или по диагонали, сомкнув или полураскрыв крылья над брюшком.
Мало что известно о способности вида расселяться. В линейной среде обитания (канава) зафиксировано, что самки значительно более подвижны, чем самцы, при этом более высокая доля самцов возвращалась примерно в одну и ту же часть канавы в последовательные дни (54,7% самцов и 38,0% самок возвращались в пределы 19 м), хотя максимальное расстояние перемещение (207 м для самцов и 192 м для самок) в последующие дни, было одинаковым.
Согласно другим исследованиям, средняя продолжительность жизни половозрелых особей, т. е. тех, кто пережил период созревания, составляет 6,8 дня для самцов и 6,6 дня для самок. Эти цифры дают общую продолжительность имаго 19,4 дня (12,6 предрепродуктивных + 6,8 репродуктивных) для самцов и 21,6 дней (15 предрепродуктивных + 6,6 репродуктивных) для самок. Вместе с тем максимальная зарегистрированная продолжительность жизни имаго - 46 дней.
Летают между растениями, растущими близко к берегам рек, между камышами и тростниками. В жаркую погоду летают быстро. В холодную погоду могут греться на светлых поверхностях, включая одежду. Отдыхают горизонтально или по диагонали, сомкнув или полураскрыв крылья над брюшком.
Мало что известно о способности вида расселяться. В линейной среде обитания (канава) зафиксировано, что самки значительно более подвижны, чем самцы, при этом более высокая доля самцов возвращалась примерно в одну и ту же часть канавы в последовательные дни (54,7% самцов и 38,0% самок возвращались в пределы 19 м), хотя максимальное расстояние перемещение (207 м для самцов и 192 м для самок) в последующие дни, было одинаковым.

♀ Фото © Семён Левый, г. Могилев
Территориальность взрослых самцов рассматривается по-разному, одни авторы указывают, что самцы очень территориальны, другие - что территориальность выражена не явно. Однако и те и другие подчеркивают их повышенную агрессивность. Самцы не только отгоняют конкурентов, но и бросаются в погоню за любыми пролетающими насекомыми. Вместе с тем, наиболее правдоподобным является предположение, что территориальность зависит от плотности популяции - чем плотность ниже, тем более очевидно проявляется территориальное поведение, и наоборот.
Самец защищает присаду, с которой он приближается к потенциальным самкам, а также к тандемным парам и патрулирующим самцам. Отмечено, что резидентные самцы выиграли 97,5% (117 из 120) территориальных споров, несмотря на то, что часто резидентный самец был меньше (в 56 столкновениях), поэтому размер, по-видимому, не влияет на успех спаривания. Перенос сперматозоидов самцом в дополнительные половые органы происходит во время тандемной фазы. Само спаривание часто происходит на листьях деревьев (например, ольхи) у кромки воды. В спокойном состоянии совокупление длится до 15 минут.
Самец защищает присаду, с которой он приближается к потенциальным самкам, а также к тандемным парам и патрулирующим самцам. Отмечено, что резидентные самцы выиграли 97,5% (117 из 120) территориальных споров, несмотря на то, что часто резидентный самец был меньше (в 56 столкновениях), поэтому размер, по-видимому, не влияет на успех спаривания. Перенос сперматозоидов самцом в дополнительные половые органы происходит во время тандемной фазы. Само спаривание часто происходит на листьях деревьев (например, ольхи) у кромки воды. В спокойном состоянии совокупление длится до 15 минут.

♂ Фото © Владимир Бондарь, Могилевский р-н
Как это типично для семейства в целом, указывается, что самец всегда охраняет самку во время яйцекладки. Он принимает вертикальное положение (сторожевую позицию) и удерживает самку, пока она опускает брюшко в воду и откладывает яйца в плавающие растения. Обычно самка постоянно погружает брюшко не дальше точки соприкосновения крыльев с поверхностью воды. Однако не всё так однозначно. Во-первых, отмечено, что иногда самки откладывают яйца в одиночку, даже в компании других пар, а свободные самцы в это время ищут себе новую пару. Это означает, что в небольшой популяции охрана контакта не всегда необходима. Во-вторых, отмечены случаи погружения самки при яйцекладке под воду, причем в некоторых случаях вместе с ней под воду погружался и самец. Один из таких случаев заснят на видео в Норфолке в 2008 г., когда самка пробыла под водой 11 минут, а самец более 4 минут. На том же видео отмечена и вторая пара, которая также погружалась под воду, хотя время погружения самки и самца во втором случае не указывается. Следует отметить, что описанные случаи погружения под воду обоих членов пары происходили при довольно высокой плотности этой локальной популяции, а в другие дни, когда плотность была ниже, яйцекладка происходила по "классической" схеме - без погружения под воду самки и, соответственно, самца.

♂ Photo © halinaa / iNaturalist.org. г. Жодино, Минская обл. CC BY-NC 4.0
В старой советской литературе указывается, что яйца откладываются партиями примерно по 350 штук. Это сопоставимо с новыми данными из Европы, однако количество яиц может значительно отличаться как локально, так и в зависимости не только от погоды, но и от сезона. Самки, откладывающие яйца в начале года, откладывают более крупные кладки из более крупных яиц, чем те, которые откладывают яйца в конце сезона, независимо от размера тела.
Скорость откладки яиц может различаться как из-за поведенческих различий между популяциями, так и из-за погодных условий. На участке в северной Германии самки откладывали яйца со скоростью 5,6±1,7 яиц в минуту, тогда как на участке в Йоркшире (Великобритания) они откладывали яйца со средней скоростью 10,8±0,4 яиц в минуту. В обоих случаях самки откладывали яйца на виды Potamogeton, и оба исследования проводились при сопоставимых температурах воды. В Германии контакт брюшка самки с листом P. natans длился от 3 с до 26,22 мин за одно посещение, но откладывание яиц во время визитов продолжительностью менее 1 мин было необычно. Средняя продолжительность ненарушенного одиночного периода яйцекладки длилась 9,9±6,6 мин, при этом самка откладывала в среднем 57,7±43,4 яиц. Однако обычно самка откладывает всю свою кладку более чем за один раз, другие авторы зафиксировали, что самки тратят в среднем 22,8±1,8 минуты на общую откладку яиц при среднем размере кладки 245 яиц (или 10,8 яиц в минуту), и подсчитали, что среднее количество кладок, отложенных самкой, составило 5,91. Это дает в общей сложности 1447 яиц, отложенных за всю ее жизнь. В этом исследовании погода была хорошей, и яйцекладка наблюдалась во все дни. Однако в сезон размножения с плохой погодой количество яиц может быть намного ниже.
Скорость откладки яиц может различаться как из-за поведенческих различий между популяциями, так и из-за погодных условий. На участке в северной Германии самки откладывали яйца со скоростью 5,6±1,7 яиц в минуту, тогда как на участке в Йоркшире (Великобритания) они откладывали яйца со средней скоростью 10,8±0,4 яиц в минуту. В обоих случаях самки откладывали яйца на виды Potamogeton, и оба исследования проводились при сопоставимых температурах воды. В Германии контакт брюшка самки с листом P. natans длился от 3 с до 26,22 мин за одно посещение, но откладывание яиц во время визитов продолжительностью менее 1 мин было необычно. Средняя продолжительность ненарушенного одиночного периода яйцекладки длилась 9,9±6,6 мин, при этом самка откладывала в среднем 57,7±43,4 яиц. Однако обычно самка откладывает всю свою кладку более чем за один раз, другие авторы зафиксировали, что самки тратят в среднем 22,8±1,8 минуты на общую откладку яиц при среднем размере кладки 245 яиц (или 10,8 яиц в минуту), и подсчитали, что среднее количество кладок, отложенных самкой, составило 5,91. Это дает в общей сложности 1447 яиц, отложенных за всю ее жизнь. В этом исследовании погода была хорошей, и яйцекладка наблюдалась во все дни. Однако в сезон размножения с плохой погодой количество яиц может быть намного ниже.

imm. Photo © halinaa / iNaturalist.org. г. Жодино, Минская обл. CC BY-NC 4.0
Яйца имеют размеры около 0,95 х 0,25 мм. Они имеют цилиндрическую форму с закругленным основанием и заостренным верхом. Когда они только что отложены, то полупрозрачны, но через несколько дней становятся бледно-коричневыми.
В ряде исследований основными местами откладки яиц были отмечены на рдесте Potamogeton natans, но откладка яиц наблюдалась и в другие водные растения, такие как P. polygonifolius, Menyanthes trifoliata, Sphagnum sp., Veronica beccabunga и иногда в согнутые в воду стебли Juncus sp. или даже плавающие остатки камыша и тростника. В P. natans большинство яиц откладываются в черешок или основание листа, но некоторые откладываются на нижнюю сторону листа. В единичных случаях используются и скопления нитчатых водорослей, между которыми затем и живут личинки.
Согласно разным экспериментам, яйца в неволе вылупляются через 18 дней, но при содержании в тени при температуре около 15°С - через 32 дня. Однако когда в лабораторию были помещены стебли P. natans, в которые самки отложили яйца, обнаружено, что партии яиц различаются как по успешности вылупления, так и по времени, которое требовалось для их вылупления. Общий процент успешного вылупления около 75%, причем более 60% яиц вылупляются в течение 24-26 дней после кладки.
В ряде исследований основными местами откладки яиц были отмечены на рдесте Potamogeton natans, но откладка яиц наблюдалась и в другие водные растения, такие как P. polygonifolius, Menyanthes trifoliata, Sphagnum sp., Veronica beccabunga и иногда в согнутые в воду стебли Juncus sp. или даже плавающие остатки камыша и тростника. В P. natans большинство яиц откладываются в черешок или основание листа, но некоторые откладываются на нижнюю сторону листа. В единичных случаях используются и скопления нитчатых водорослей, между которыми затем и живут личинки.
Согласно разным экспериментам, яйца в неволе вылупляются через 18 дней, но при содержании в тени при температуре около 15°С - через 32 дня. Однако когда в лабораторию были помещены стебли P. natans, в которые самки отложили яйца, обнаружено, что партии яиц различаются как по успешности вылупления, так и по времени, которое требовалось для их вылупления. Общий процент успешного вылупления около 75%, причем более 60% яиц вылупляются в течение 24-26 дней после кладки.

♂ Photo © Станислав Бакей / iNaturalist.org. Новогрудский р-н, Гродненская обл. CC BY-NC 4.0
Нимфа. Окраска от темно-желтоватой до коричневой с дорсальной стороны, более светлая с вентральной. Тело относительно короткое, длиной 17-19 мм, шириной 3,5-4 мм. Голова относительно большая, широкая, около 3,25 мм в поперечнике, впереди округлая; на темени глазки ясно видимы; глаза большие, выдающиеся; членики антенн посредине темные; затылок относительно узкий, отделен от глаз волнообразной линией, боковые края значительно скошены назад; задние углы тупо закругленные, задний край сильно вогнутый; позади глаз 2 изогнутые светлые полоски, по середине затылка 2 светлых прямоугольных с темными гранями пятна. Маска в состоянии покоя доходит до основания передней пары ног; подбородок плоский, пятиугольный, узкий у основания, широкий у вершины. Переднегрудь короткая, узкая, но шире и немного выше, чем затылок, с 4 округлыми углами, со светлыми отметинами. Крыловые чехлы прямые, длинные, достигают почти дистального края IV сегмента брюшка. Ноги довольно длинные, светлее основной окраски тела, бедра; с 2 темными кольцами, голени с 1 более светлым кольцом; передние ноги длиной 7 мм, средние 8 мм, задние 11 мм. Брюшко цилиндрическое, постепенно суживающееся к концу, со светлой срединной дорсальной, продольной полосой, проходящей по всем сегментам, и светлой боковой линией вдоль каждого сегмента, особенно развитой у старых особей; на каждом сегменте, за исключением Х, по 2 темных точки, расположенных с каждой стороны светлой срединной дорсальной полосы; Х сегмент по бокам с короткими шипиками, назади с срединной выемкой: межсегментальные пространства с несколькими маленькими белыми точками. Хвостовые жаберные пластинки довольно короткие, длиной 5-5,5 мм, шириной 2-2,5 мм, эллипсоидальной формы, очень узкие у основания и сильно расширяющиеся к заднему концу, с небольшим срединным острием; срединный трахейный ствол ясный; разветвления трахеи без специального просветления почти не видно; пластинки светлые, с большим поперечным темным пятном посредине и многими маленькими темными пятнышками. Створки яйцеклада самки простираются до дистального края Х стернита, на вершине заостренные.

♂ Фото © Денис Китель, Россонский р-н (Витебская обл.)
Это один из немногих известных территориальных видов на личиночной стадии. Успешное территориальное поведение приводит к более крупным личинкам и, следовательно, к более успешному размножению взрослых особей. Личинки, как правило, живут на дне или вблизи дна среди водной растительности или детрита. Они обладают одной из самых широких экологических устойчивостей, приспособлены к широкому диапазону pH и менее чувствительны к эвтрофикации.
Обнаружено, что некоторые из только что вылупившихся личинок в месте откладки яиц (Potamogeton natans) пассивно рассеивались водными потоками от места откладки яиц, но передвижение более старых личинок было незначительным, хотя некоторое передвижение вглубь водоема происходило на второе лето. Согласно другому исследованию, только что вылупившиеся личинки, как правило, оставались рядом с местами откладки яиц (Potamogeton) в течение июля и августа, но в сентябре перемещались в район Juncus spp., где зимовали. Однако они, как правило, остаются на одном и том же месте в течение второго года развития. Таким образом, в этом последнем исследовании имело место почти полное пространственное разделение годичных классов, что может иметь важное значение для предотвращения хищничества более мелких особей более крупными. Ряд исследований показал, что количество личинок, пойманных сеткой, уменьшается зимой (так называемое «зимнее исчезновение»), а весной снова увеличивается.
Личинки поедают разнообразную добычу, их основной пищей являются хирономиды и низшие ракообразные, но также отлавливаются личинки подёнок Leptophlebia spp., планарии и, возможно, другие личинки P. nymphula. При анализе экскрементов личинок от 4-го до последнего (12-го) возраста обнаружено, что личинки хирономид дают 60-75% потребляемой пищевой энергии. В лабораторном эксперименте с использованием дафний в качестве добычи обнаружено, что эффективность ассимиляции снижается с увеличением размера личинок примерно с 95% во втором возрасте до примерно 86% в последнем возрасте. Личинки не демонстрируют какой-либо метаболической акклиматизации, их частота дыхания увеличивается с повышением температуры. Подсчитано, что 42-43% потребляемой личинками энергии расходуется на рост и дыхание.
Основными хищниками личинок, вероятно, являются рыбы. В лаборатории также было замечено, что личинки ручейника Phryganea поедают личинок P. nymphula. У молодых возрастов уровень смертности выше, чем у более поздних, поскольку их небольшой размер делает их уязвимыми для более широкого круга хищников, включая более крупных особей своего же вида.
Обнаружено, что некоторые из только что вылупившихся личинок в месте откладки яиц (Potamogeton natans) пассивно рассеивались водными потоками от места откладки яиц, но передвижение более старых личинок было незначительным, хотя некоторое передвижение вглубь водоема происходило на второе лето. Согласно другому исследованию, только что вылупившиеся личинки, как правило, оставались рядом с местами откладки яиц (Potamogeton) в течение июля и августа, но в сентябре перемещались в район Juncus spp., где зимовали. Однако они, как правило, остаются на одном и том же месте в течение второго года развития. Таким образом, в этом последнем исследовании имело место почти полное пространственное разделение годичных классов, что может иметь важное значение для предотвращения хищничества более мелких особей более крупными. Ряд исследований показал, что количество личинок, пойманных сеткой, уменьшается зимой (так называемое «зимнее исчезновение»), а весной снова увеличивается.
Личинки поедают разнообразную добычу, их основной пищей являются хирономиды и низшие ракообразные, но также отлавливаются личинки подёнок Leptophlebia spp., планарии и, возможно, другие личинки P. nymphula. При анализе экскрементов личинок от 4-го до последнего (12-го) возраста обнаружено, что личинки хирономид дают 60-75% потребляемой пищевой энергии. В лабораторном эксперименте с использованием дафний в качестве добычи обнаружено, что эффективность ассимиляции снижается с увеличением размера личинок примерно с 95% во втором возрасте до примерно 86% в последнем возрасте. Личинки не демонстрируют какой-либо метаболической акклиматизации, их частота дыхания увеличивается с повышением температуры. Подсчитано, что 42-43% потребляемой личинками энергии расходуется на рост и дыхание.
Основными хищниками личинок, вероятно, являются рыбы. В лаборатории также было замечено, что личинки ручейника Phryganea поедают личинок P. nymphula. У молодых возрастов уровень смертности выше, чем у более поздних, поскольку их небольшой размер делает их уязвимыми для более широкого круга хищников, включая более крупных особей своего же вида.

♂ Фото © Семён Левый, Борисовский р-н, Минская обл.
Развитие обычно семивольтинное. Личинки обычно развиваются 2 года, но развитие может занять 1 год или, а при высокой плотности личинок, 3 года.
Включая короткую предличиночную стадию, у вида насчитывается 12 личиночных возрастов. Как правило, личинки зимуют сначала в ряде ранних возрастов (чаще 5-го и 6-го) и переходят в предпоследний возраст на второе лето, оставаясь в этом возрасте до конца августа, после чего быстро переходят в последний возраст в течение следующего месяца и зимуют в этом возрасте. Задержка старшей возрастной группы в предпоследнем возрасте летом, а затем в последнем зимой уменьшает временную изменчивость среди личинок и в значительной степени способствует синхронизации метаморфоза с последующим вылетом в начале лета (май-начало июля). Переменная, обычно небольшая, часть когорты проводит вторую зиму в предпоследнем возрасте. Весной такие личинки быстро входят в фазу последнего возраста в мае и приступают непосредственно к метаморфозу и вылету. Такие особи мельче и во взрослом состоянии, чем те, которые перезимовали в последнем возрасте, и они составляют второй, более поздний компонент формирующейся популяции.
Однако при исключительно большом количестве молодых личинок некоторым требуется 3 года для полного развития, не достигая предпоследней стадии к началу второй зимы, возможно, из-за конкуренции за пищу. Предполагается, что такие личинки, вероятно, имеются в большинстве лет. Это связано с тем, что для всех личинок может не хватать хороших кормовых участков, особенно в годы их высокой численности. Личинки, занимающие места, где пища находится в пределах досягаемости, растут быстро, а личинки в субоптимальных местах, где пищи мало, растут медленнее; последние вполне могут погибнуть, если не появятся лучшие места для кормления.
Включая короткую предличиночную стадию, у вида насчитывается 12 личиночных возрастов. Как правило, личинки зимуют сначала в ряде ранних возрастов (чаще 5-го и 6-го) и переходят в предпоследний возраст на второе лето, оставаясь в этом возрасте до конца августа, после чего быстро переходят в последний возраст в течение следующего месяца и зимуют в этом возрасте. Задержка старшей возрастной группы в предпоследнем возрасте летом, а затем в последнем зимой уменьшает временную изменчивость среди личинок и в значительной степени способствует синхронизации метаморфоза с последующим вылетом в начале лета (май-начало июля). Переменная, обычно небольшая, часть когорты проводит вторую зиму в предпоследнем возрасте. Весной такие личинки быстро входят в фазу последнего возраста в мае и приступают непосредственно к метаморфозу и вылету. Такие особи мельче и во взрослом состоянии, чем те, которые перезимовали в последнем возрасте, и они составляют второй, более поздний компонент формирующейся популяции.
Однако при исключительно большом количестве молодых личинок некоторым требуется 3 года для полного развития, не достигая предпоследней стадии к началу второй зимы, возможно, из-за конкуренции за пищу. Предполагается, что такие личинки, вероятно, имеются в большинстве лет. Это связано с тем, что для всех личинок может не хватать хороших кормовых участков, особенно в годы их высокой численности. Личинки, занимающие места, где пища находится в пределах досягаемости, растут быстро, а личинки в субоптимальных местах, где пищи мало, растут медленнее; последние вполне могут погибнуть, если не появятся лучшие места для кормления.

♂ Photo © kukarzinio / iNaturalist.org. Лидский р-н, Гродненская обл. CC BY-NC 4.0
Окрыление имаго происходит синхронно и вид является одной из первых стрекоз, летающих с конца апреля. Начало вылета колеблется от года к году, чем раньше температура воды достигает 10-12°С, тем раньше начало отрождений. Летом происходит второе, меньшее по размеру, несинхронизированное появление имаго, и небольшое количество взрослых особей могут летать до начала сентября.
Отрождение начинается примерно в 07:30, причем пик обычно приходится на период с 08:00 до 09:00. Однако при неблагоприятной погоде отрождение переносится на следующий день. Регистрации о высоте, на которой происходит отрождение имаго значительно различаются. В зависимости от местности они могут быть от нескольких сантиметров над уровнем воды (85-90%) на травянистых растениях до 1,5 м на стволах деревьев. Особи требуется 1 или 2 часа, чтобы завершить свой метаформоз.
Данные о выживании имаго в период предварительного созревания получить сложно, поскольку на этом этапе стрекозы покидают водоем и, следовательно, их труднее контролировать. Тем не менее, показатель выживаемости в этот период, по крайней мере, такой же, как и у зрелых особей. Подсчитано, что стрекозы возвращаются к водоему примерно через 15 дней после отрождения, в течение которых они достигают половой зрелости. Самкам требуется больше времени (в среднем 18,6 дня) для созревания, чем самцам (в среднем 12,6 дня). Однако эти цифры основаны на первом зарегистрированном посещении водоема. Поскольку некоторые особи, возможно, не были замечены во время их первого посещения, эти цифры почти наверняка завышены, что особенно относится к самкам, поскольку между посещениями водоема они проводят больше времени вдали от воды, поэтому цифра в 15 дней для самок может быть более точной. Кроме того, имаго редко можно увидеть летающими, когда не светит солнце.
Отрождение начинается примерно в 07:30, причем пик обычно приходится на период с 08:00 до 09:00. Однако при неблагоприятной погоде отрождение переносится на следующий день. Регистрации о высоте, на которой происходит отрождение имаго значительно различаются. В зависимости от местности они могут быть от нескольких сантиметров над уровнем воды (85-90%) на травянистых растениях до 1,5 м на стволах деревьев. Особи требуется 1 или 2 часа, чтобы завершить свой метаформоз.
Данные о выживании имаго в период предварительного созревания получить сложно, поскольку на этом этапе стрекозы покидают водоем и, следовательно, их труднее контролировать. Тем не менее, показатель выживаемости в этот период, по крайней мере, такой же, как и у зрелых особей. Подсчитано, что стрекозы возвращаются к водоему примерно через 15 дней после отрождения, в течение которых они достигают половой зрелости. Самкам требуется больше времени (в среднем 18,6 дня) для созревания, чем самцам (в среднем 12,6 дня). Однако эти цифры основаны на первом зарегистрированном посещении водоема. Поскольку некоторые особи, возможно, не были замечены во время их первого посещения, эти цифры почти наверняка завышены, что особенно относится к самкам, поскольку между посещениями водоема они проводят больше времени вдали от воды, поэтому цифра в 15 дней для самок может быть более точной. Кроме того, имаго редко можно увидеть летающими, когда не светит солнце.

Photo © kukarzinio / iNaturalist.org. Лидский р-н, Гродненская обл. CC BY-NC 4.0

Göran Liljeberg & Hallvard Elven. Naturhistorisk museum, Universitetet i Oslo. CC BY-SA 3.0

♂ Image © Göran Liljeberg / Naturforskaren. CC BY 3.0
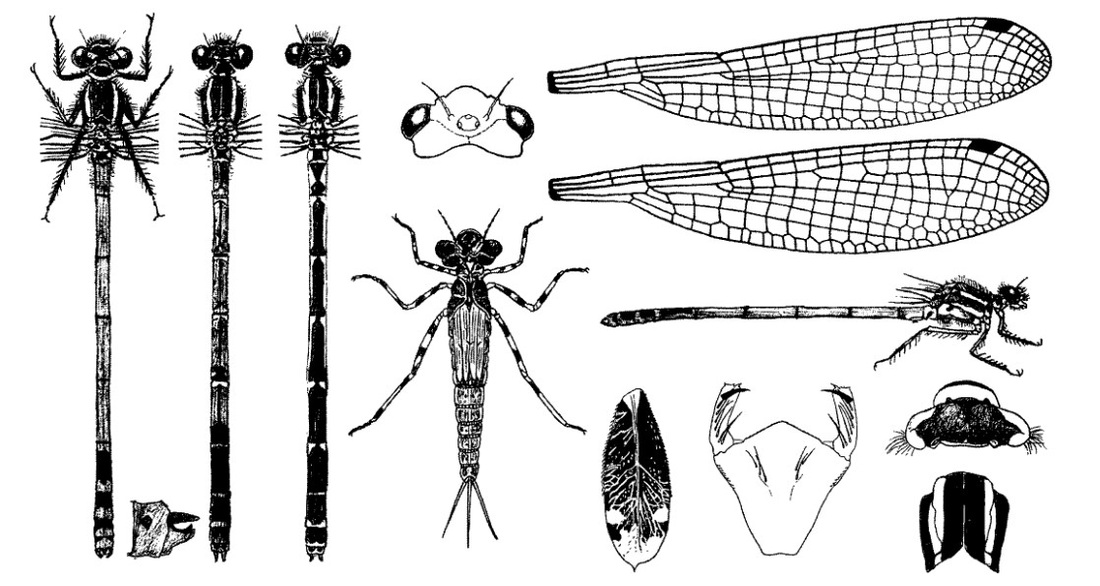
Скворцов В. Э. 2010. Стрекозы Восточной Европы и Кавказа: Атлас-определитель.
Источники
1. Спурис З. Д. "Отряд Odonatoptera - Стрекозы" / Бей-Биенко Г. Я. (ред.) "Определитель насекомых европейской части СССР". т. I, Москва - Ленинград, 1964. С. 137-162.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Large Red Damsel. Pyrrhosoma nymphula" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 50-51.
9. Smallshire D., Swash A. "Large Red Damselfly. Pyrrhosoma nymphula" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 72-73.
10. Moreno-Benítez J. M., Ripoll J. "Pyrrhosoma nymphula" / in: Dragonflies along the Great Málaga Path and in the Province. Identification Guide, 2018. pp. 86-87.
11. Corbet P. S., Harvey I. F. "Seasonal regulation in Pyrrhosoma nymphula (Sulzer) (Zygoptera: Coenagrionidae). 1. Seasonal development in nature" / Odonalologica 18(2): 133-145. 1989.
12. Mill P. J. "Species Review 3: The Large Red Damselfly Pyrrhosoma nymphula (Sulzer) with notes on its close relative the Greek Red Damselfly Pyrrhosoma elisabethae Schmidt" / Journal of the British Dragonfly Society, Volume 26 No. 1, 2010. pp. 34-56.
13. Shwerin G. "Submergence of both sexes during oviposition in the Large Red Damselfly Pyrrhosoma nymphula (Sulzer) in Norfolk" / Journal of the British Dragonfly Society, Volume 25 No. 2, 2009. pp. 62-67.
14. Lehmann A., Lehmann G., Mauersberger R. "Frühe Adonislibelle – Pyrrhosoma nymphula (Sulzer 1776)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 32-33.
2. Скворцов В. Э. "Стрекозы Восточной Европы и Кавказа: Атлас-определитель". Москва, 2010. - 623 с.
3. Татаринов А. Г., Кулакова О. И. "Стрекозы" (Фауна европейского Северо-Востока России. т. Х). Санкт-Петербург, 2009. - 213 с.
4. Попова А. Н. "Личинки стрекоз фауны СССР (Odonata)". Москва-Ленинград, 1950. - 235 с.
5. Дьяконов А. М. "Наши стрекозы. Определитель стрекоз и их личинок". Ленинград, 1926. - 72 с.
6. Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. "Определитель насекомых европейской части СССР". Москва, 1972. - 304 с.
7. Merritt R., Moore N. W., Eversham B. C. "Atlas of the dragonflies of Britain and Ireland". ITE research publication no. 9. Centre for Ecology and Hydrology. Natural Environment Research Council. London, 1996. - 149 pp.
8. Smallshire D., Swash A. "Large Red Damsel. Pyrrhosoma nymphula" / Europe's Dragonflies: A field guide to the damselflies and dragonflies. Princeton University Press, 2020. pp. 50-51.
9. Smallshire D., Swash A. "Large Red Damselfly. Pyrrhosoma nymphula" / Britain's Dragonflies. A Field Guide to the Damselflies and Dragonflies of Great Britain and Ireland. Fully Revised and Updated Third Edition. Princeton University Press, 2014. pp. 72-73.
10. Moreno-Benítez J. M., Ripoll J. "Pyrrhosoma nymphula" / in: Dragonflies along the Great Málaga Path and in the Province. Identification Guide, 2018. pp. 86-87.
11. Corbet P. S., Harvey I. F. "Seasonal regulation in Pyrrhosoma nymphula (Sulzer) (Zygoptera: Coenagrionidae). 1. Seasonal development in nature" / Odonalologica 18(2): 133-145. 1989.
12. Mill P. J. "Species Review 3: The Large Red Damselfly Pyrrhosoma nymphula (Sulzer) with notes on its close relative the Greek Red Damselfly Pyrrhosoma elisabethae Schmidt" / Journal of the British Dragonfly Society, Volume 26 No. 1, 2010. pp. 34-56.
13. Shwerin G. "Submergence of both sexes during oviposition in the Large Red Damselfly Pyrrhosoma nymphula (Sulzer) in Norfolk" / Journal of the British Dragonfly Society, Volume 25 No. 2, 2009. pp. 62-67.
14. Lehmann A., Lehmann G., Mauersberger R. "Frühe Adonislibelle – Pyrrhosoma nymphula (Sulzer 1776)" / im: Mauersberger R. et al. Die Libellenfauna des Landes Brandenburg. Natursch. Landschaftspfl. Bbg., 2013, 22 (3, 4): 32-33.
